28.7 Patterns of Inheritance
Learning Objectives
By the end of this section, you will be able to:
- Differentiate between genotype and phenotype
- Describe how alleles determine a person’s traits
- Summarize Mendel’s experiments and relate them to human genetics
- Explain the inheritance of autosomal dominant and recessive and sex-linked genetic disorders
We have discussed the events that lead to the development of a newborn. But what makes each newborn unique? The answer lies, of course, in the DNA in the sperm and oocyte that combined to produce that first diploid cell, the human zygote.
From Genotype to Phenotype
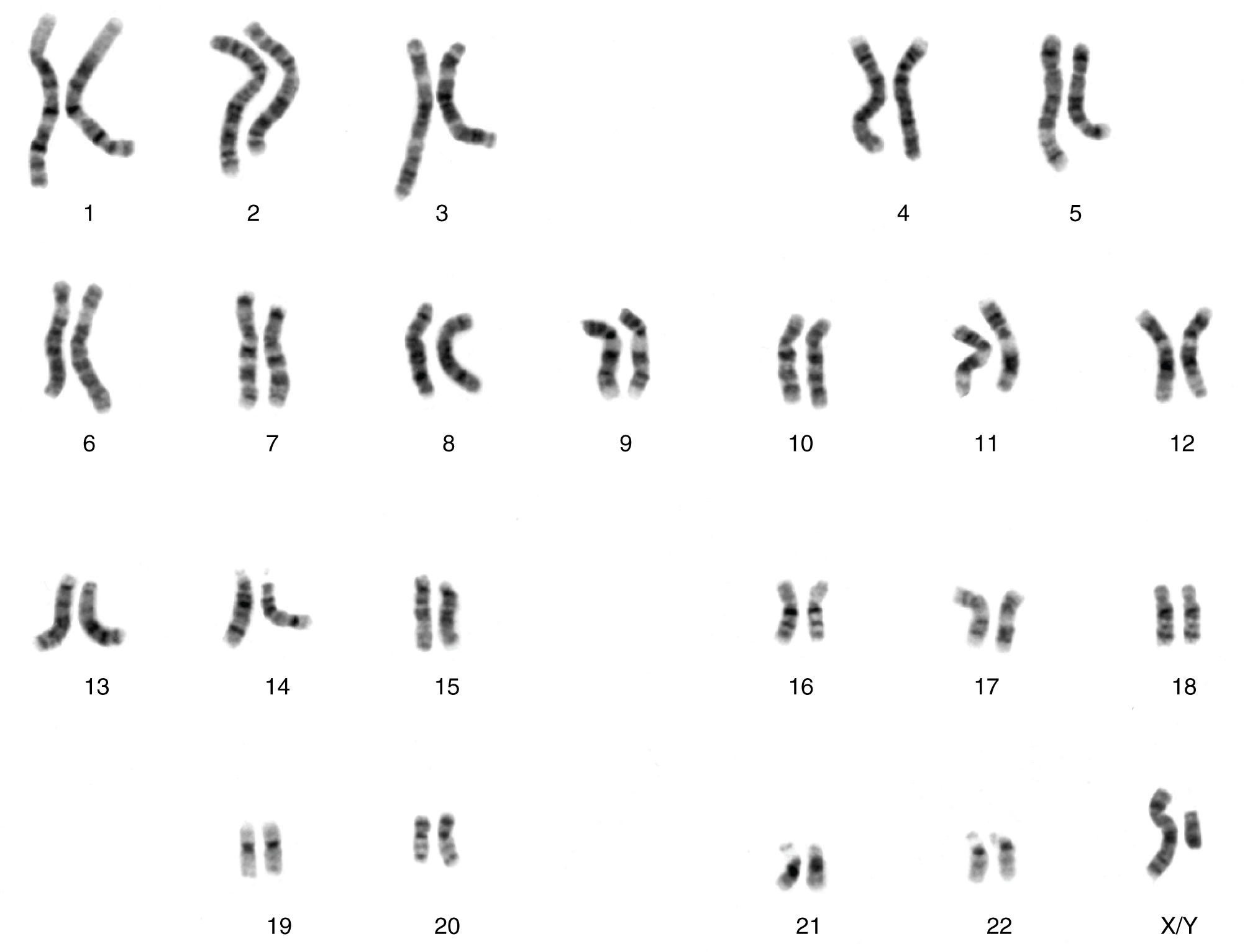
Each human body cell has a full complement of DNA stored in 23 pairs of chromosomes. Figure 28.7.1 shows the pairs in a systematic arrangement called a karyotype. Among these is one pair of chromosomes, called the sex chromosomes, that determines the sex of the individual (XX in females, XY in males). The remaining 22 chromosome pairs are called autosomal chromosomes. Each of these chromosomes carries hundreds or even thousands of genes, each of which codes for the assembly of a particular protein—that is, genes are “expressed” as proteins. An individual’s complete genetic makeup is referred to as his or her genotype. The characteristics that the genes express, whether they are physical, behavioral, or biochemical, are a person’s phenotype.
You inherit one chromosome in each pair—a full complement of 23—from each parent. This occurs when the sperm and oocyte combine at the moment of your conception. Homologous chromosomes—those that make up a complementary pair—have genes for the same characteristics in the same location on the chromosome. Because one copy of a gene, an allele, is inherited from each parent, the alleles in these complementary pairs may vary. Take for example an allele that encodes for dimples. A child may inherit the allele encoding for dimples on the chromosome from the father and the allele that encodes for smooth skin (no dimples) on the chromosome from the mother.
Although a person can have two identical alleles for a single gene (a homozygous state), it is also possible for a person to have two different alleles (a heterozygous state). The two alleles can interact in several different ways. The expression of an allele can be dominant, for which the activity of this gene will mask the expression of a nondominant, or recessive, allele. Sometimes dominance is complete; at other times, it is incomplete. In some cases, both alleles are expressed at the same time in a form of expression known as codominance.
In the simplest scenario, a single pair of genes will determine a single heritable characteristic. However, it is quite common for multiple genes to interact to confer a feature. For instance, eight or more genes—each with their own alleles—determine eye color in humans. Moreover, although any one person can only have two alleles corresponding to a given gene, more than two alleles commonly exist in a population. This phenomenon is called multiple alleles. For example, there are three different alleles that encode ABO blood type; these are designated IA, IB, and i.
Over 100 years of theoretical and experimental genetics studies, and the more recent sequencing and annotation of the human genome, have helped scientists to develop a better understanding of how an individual’s genotype is expressed as their phenotype. This body of knowledge can help scientists and medical professionals to predict, or at least estimate, some of the features that an offspring will inherit by examining the genotypes or phenotypes of the parents. One important application of this knowledge is to identify an individual’s risk for certain heritable genetic disorders. However, most diseases have a multigenic pattern of inheritance and can also be affected by the environment, so examining the genotypes or phenotypes of a person’s parents will provide only limited information about the risk of inheriting a disease. Only for a handful of single-gene disorders can genetic testing allow clinicians to calculate the probability with which a child born to the two parents tested may inherit a specific disease.
Mendel’s Theory of Inheritance
Our contemporary understanding of genetics rests on the work of a nineteenth-century monk. Working in the mid-1800s, long before anyone knew about genes or chromosomes, Gregor Mendel discovered that garden peas transmit their physical characteristics to subsequent generations in a discrete and predictable fashion. When he mated, or crossed, two pure-breeding pea plants that differed by a certain characteristic, the first-generation offspring all looked like one of the parents. For instance, when he crossed tall and dwarf pure-breeding pea plants, all of the offspring were tall. Mendel called tallness dominant because it was expressed in offspring when it was present in a purebred parent. He called dwarfism recessive because it was masked in the offspring if one of the purebred parents possessed the dominant characteristic. Note that tallness and dwarfism are variations on the characteristic of height. Mendel called such a variation a trait. We now know that these traits are the expression of different alleles of the gene encoding height.
Mendel performed thousands of crosses in pea plants with differing traits for a variety of characteristics. And he repeatedly came up with the same results—among the traits he studied, one was always dominant, and the other was always recessive. (Remember, however, that this dominant–recessive relationship between alleles is not always the case; some alleles are codominant, and sometimes dominance is incomplete.)
Using his understanding of dominant and recessive traits, Mendel tested whether a recessive trait could be lost altogether in a pea lineage or whether it would resurface in a later generation. By crossing the second-generation offspring of purebred parents with each other, he showed that the latter was true: recessive traits reappeared in third-generation plants in a ratio of 3:1 (three offspring having the dominant trait and one having the recessive trait). Mendel then proposed that characteristics such as height were determined by heritable “factors” that were transmitted, one from each parent, and inherited in pairs by offspring.
In the language of genetics, Mendel’s theory applied to humans says that if an individual receives two dominant alleles, one from each parent, the individual’s phenotype will express the dominant trait. If an individual receives two recessive alleles, then the recessive trait will be expressed in the phenotype. Individuals who have two identical alleles for a given gene, whether dominant or recessive, are said to be homozygous for that gene (homo- = “same”). Conversely, an individual who has one dominant allele and one recessive allele is said to be heterozygous for that gene (hetero- = “different” or “other”). In this case, the dominant trait will be expressed, and the individual will be phenotypically identical to an individual who possesses two dominant alleles for the trait.
It is common practice in genetics to use capital and lowercase letters to represent dominant and recessive alleles. Using Mendel’s pea plants as an example, if a tall pea plant is homozygous, it will possess two tall alleles (TT). A dwarf pea plant must be homozygous because its dwarfism can only be expressed when two recessive alleles are present (tt). A heterozygous pea plant (Tt) would be tall and phenotypically indistinguishable from a tall homozygous pea plant because of the dominant tall allele. Mendel deduced that a 3:1 ratio of dominant to recessive would be produced by the random segregation of heritable factors (genes) when crossing two heterozygous pea plants. In other words, for any given gene, parents are equally likely to pass down either one of their alleles to their offspring in a haploid gamete, and the result will be expressed in a dominant–recessive pattern if both parents are heterozygous for the trait.
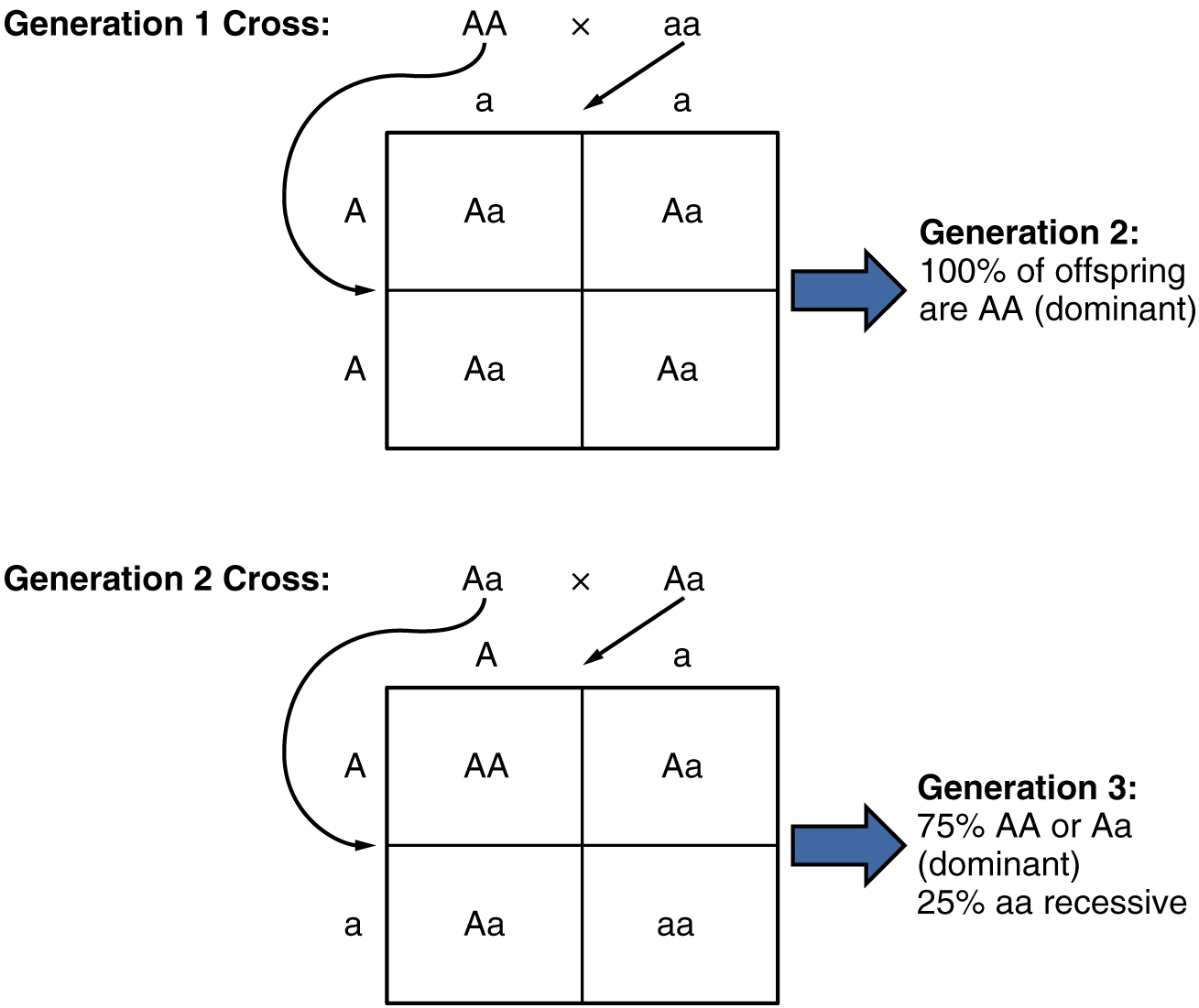
Because of the random segregation of gametes, the laws of chance and probability come into play when predicting the likelihood of a given phenotype. Consider a cross between an individual with two dominant alleles for a trait (AA) and an individual with two recessive alleles for the same trait (aa). All of the parental gametes from the dominant individual would be A, and all of the parental gametes from the recessive individual would be a (Figure 28.7.2). All of the offspring of that second generation, inheriting one allele from each parent, would have the genotype Aa, and the probability of expressing the phenotype of the dominant allele would be 4 out of 4, or 100 percent.
This seems simple enough, but the inheritance pattern gets interesting when the second-generation Aa individuals are crossed. In this generation, 50 percent of each parent’s gametes are A and the other 50 percent are a. By Mendel’s principle of random segregation, the possible combinations of gametes that the offspring can receive are AA, Aa, aA (which is the same as Aa), and aa. Because segregation and fertilization are random, each offspring has a 25 percent chance of receiving any of these combinations. Therefore, if an Aa × Aa cross were performed 1000 times, approximately 250 (25 percent) of the offspring would be AA; 500 (50 percent) would be Aa (that is, Aa plus aA); and 250 (25 percent) would be aa. The genotypic ratio for this inheritance pattern is 1:2:1. However, we have already established that AA and Aa (and aA) individuals all express the dominant trait (i.e., share the same phenotype), and can therefore be combined into one group. The result is Mendel’s third-generation phenotype ratio of 3:1.
Mendel’s observation of pea plants also included many crosses that involved multiple traits, which prompted him to formulate the principle of independent assortment. The law states that the members of one pair of genes (alleles) from a parent will sort independently from other pairs of genes during the formation of gametes. Applied to pea plants, that means that the alleles associated with the different traits of the plant, such as color, height, or seed type, will sort independently of one another. This holds true except when two alleles happen to be located close to one other on the same chromosome. Independent assortment provides for a great degree of diversity in offspring.
Mendelian genetics represent the fundamentals of inheritance, but there are two important qualifiers to consider when applying Mendel’s findings to inheritance studies in humans. First, as we’ve already noted, not all genes are inherited in a dominant–recessive pattern. Although all diploid individuals have two alleles for every gene, allele pairs may interact to create several types of inheritance patterns, including incomplete dominance and codominance.
Secondly, Mendel performed his studies using thousands of pea plants. He was able to identify a 3:1 phenotypic ratio in second-generation offspring because his large sample size overcame the influence of variability resulting from chance. In contrast, no human couple has ever had thousands of children. If we know that a man and woman are both heterozygous for a recessive genetic disorder, we would predict that one in every four of their children would be affected by the disease. In real life, however, the influence of chance could change that ratio significantly. For example, if a man and a woman are both heterozygous for cystic fibrosis, a recessive genetic disorder that is expressed only when the individual has two defective alleles, we would expect one in four of their children to have cystic fibrosis. However, it is entirely possible for them to have seven children, none of whom is affected, or for them to have two children, both of whom are affected. For each individual child, the presence or absence of a single gene disorder depends on which alleles that child inherits from his or her parents.
Autosomal Dominant Inheritance
In the case of cystic fibrosis, the disorder is recessive to the normal phenotype. However, a genetic abnormality may be dominant to the normal phenotype. When the dominant allele is located on one of the 22 pairs of autosomes (non-sex chromosomes), we refer to its inheritance pattern as autosomal dominant. An example of an autosomal dominant disorder is neurofibromatosis type I, a disease that induces tumor formation within the nervous system that leads to skin and skeletal deformities. Consider a couple in which one parent is heterozygous for this disorder (and who therefore has neurofibromatosis), Nn, and one parent is homozygous for the normal gene, nn. The heterozygous parent would have a 50 percent chance of passing the dominant allele for this disorder to his or her offspring, and the homozygous parent would always pass the normal allele. Therefore, four possible offspring genotypes are equally likely to occur: Nn, Nn, nn, and nn. That is, every child of this couple would have a 50 percent chance of inheriting neurofibromatosis. This inheritance pattern is shown in Figure 28.7.3, in a form called a Punnett square, named after its creator, the British geneticist Reginald Punnett.
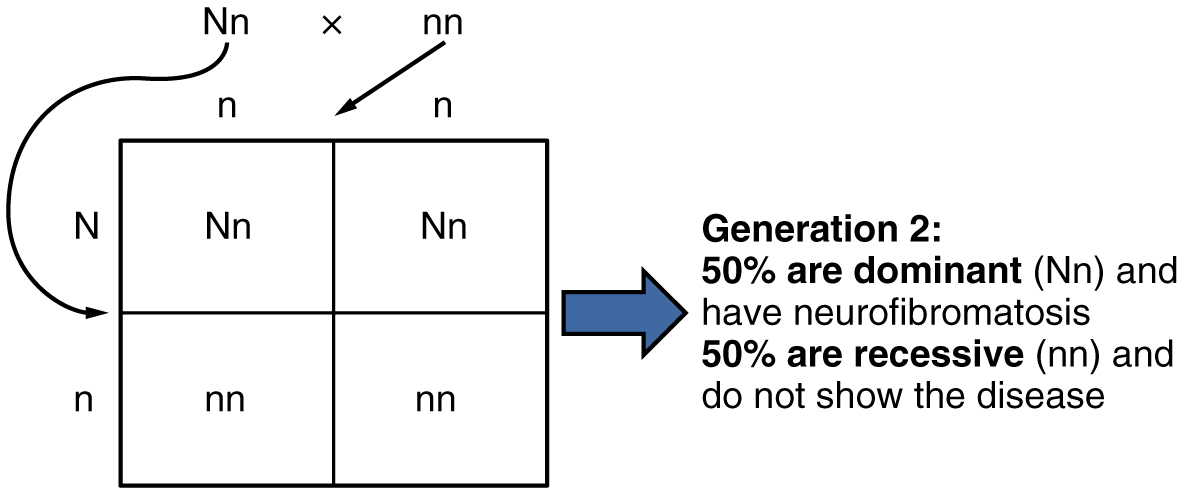
Other genetic diseases that are inherited in this pattern are achondroplastic dwarfism, Marfan syndrome, and Huntington’s disease. Because autosomal dominant disorders are expressed by the presence of just one gene, an individual with the disorder will know that he or she has at least one faulty gene. The expression of the disease may manifest later in life, after the childbearing years, which is the case in Huntington’s disease (discussed in more detail later in this section).
Autosomal Recessive Inheritance
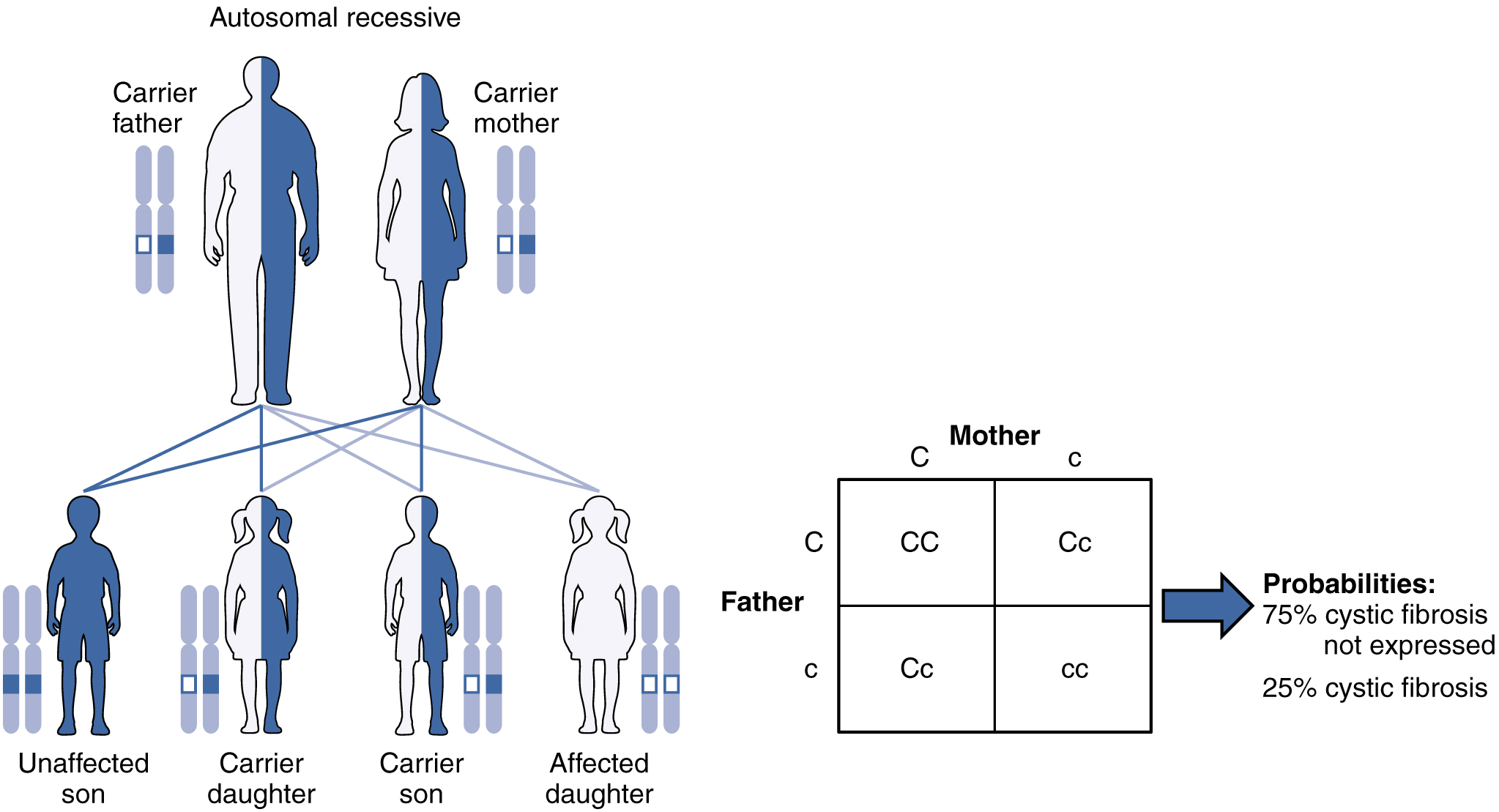
When a genetic disorder is inherited in an autosomal recessive pattern, the disorder corresponds to the recessive phenotype. Heterozygous individuals will not display symptoms of this disorder, because their unaffected gene will compensate. Such an individual is called a carrier. Carriers for an autosomal recessive disorder may never know their genotype unless they have a child with the disorder.
An example of an autosomal recessive disorder is cystic fibrosis (CF), which we introduced earlier. CF is characterized by the chronic accumulation of a thick, tenacious mucus in the lungs and digestive tract. Decades ago, children with CF rarely lived to adulthood. With advances in medical technology, the average lifespan in developed countries has increased into middle adulthood. CF is a relatively common disorder that occurs in approximately 1 in 2000 Caucasians. A child born to two CF carriers would have a 25 percent chance of inheriting the disease. This is the same 3:1 dominant:recessive ratio that Mendel observed in his pea plants would apply here. The pattern is shown in Figure 28.7.4, using a diagram that tracks the likely incidence of an autosomal recessive disorder on the basis of parental genotypes.
On the other hand, a child born to a CF carrier and someone with two unaffected alleles would have a 0 percent probability of inheriting CF, but would have a 50 percent chance of being a carrier. Other examples of autosome recessive genetic illnesses include the blood disorder sickle-cell anemia, the fatal neurological disorder Tay–Sachs disease, and the metabolic disorder phenylketonuria.
X-linked Dominant or Recessive Inheritance
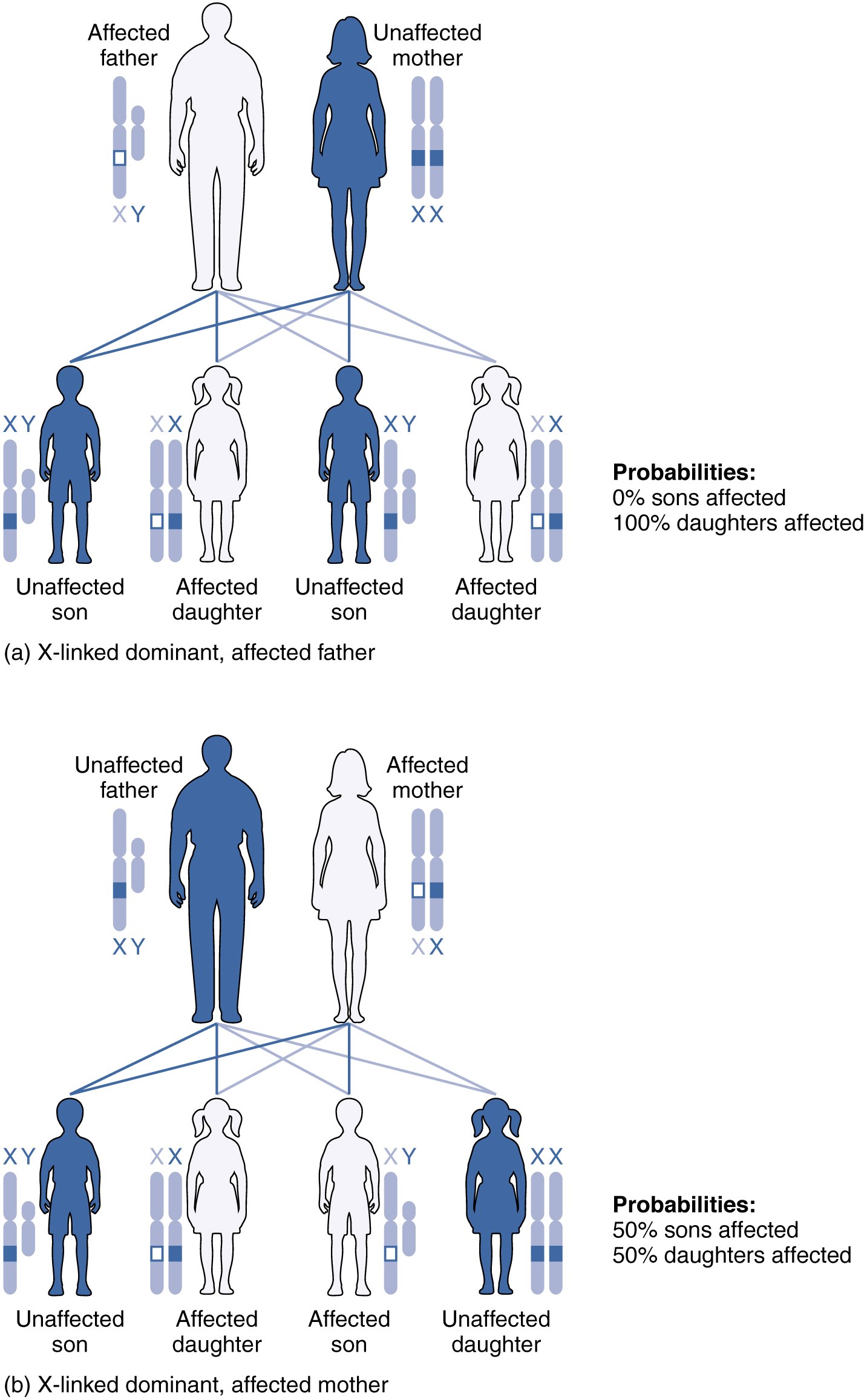
An X-linked transmission pattern involves genes located on the X chromosome of the 23rd pair (Figure 28.7.5). Recall that a male has one X and one Y chromosome. When a father transmits a Y chromosome, the child is male, and when he transmits an X chromosome, the child is female. A mother can transmit only an X chromosome, as both her sex chromosomes are X chromosomes.
When an abnormal allele for a gene that occurs on the X chromosome is dominant over the normal allele, the pattern is described as X-linked dominant. This is the case with vitamin D–resistant rickets: an affected father would pass the disease gene to all of his daughters, but none of his sons, because he donates only the Y chromosome to his sons (see Figure 28.7.5a). If it is the mother who is affected, all of her children—male or female—would have a 50 percent chance of inheriting the disorder because she can only pass an X chromosome on to her children (see Figure 28.7.5b). For an affected female, the inheritance pattern would be identical to that of an autosomal dominant inheritance pattern in which one parent is heterozygous and the other is homozygous for the normal gene.
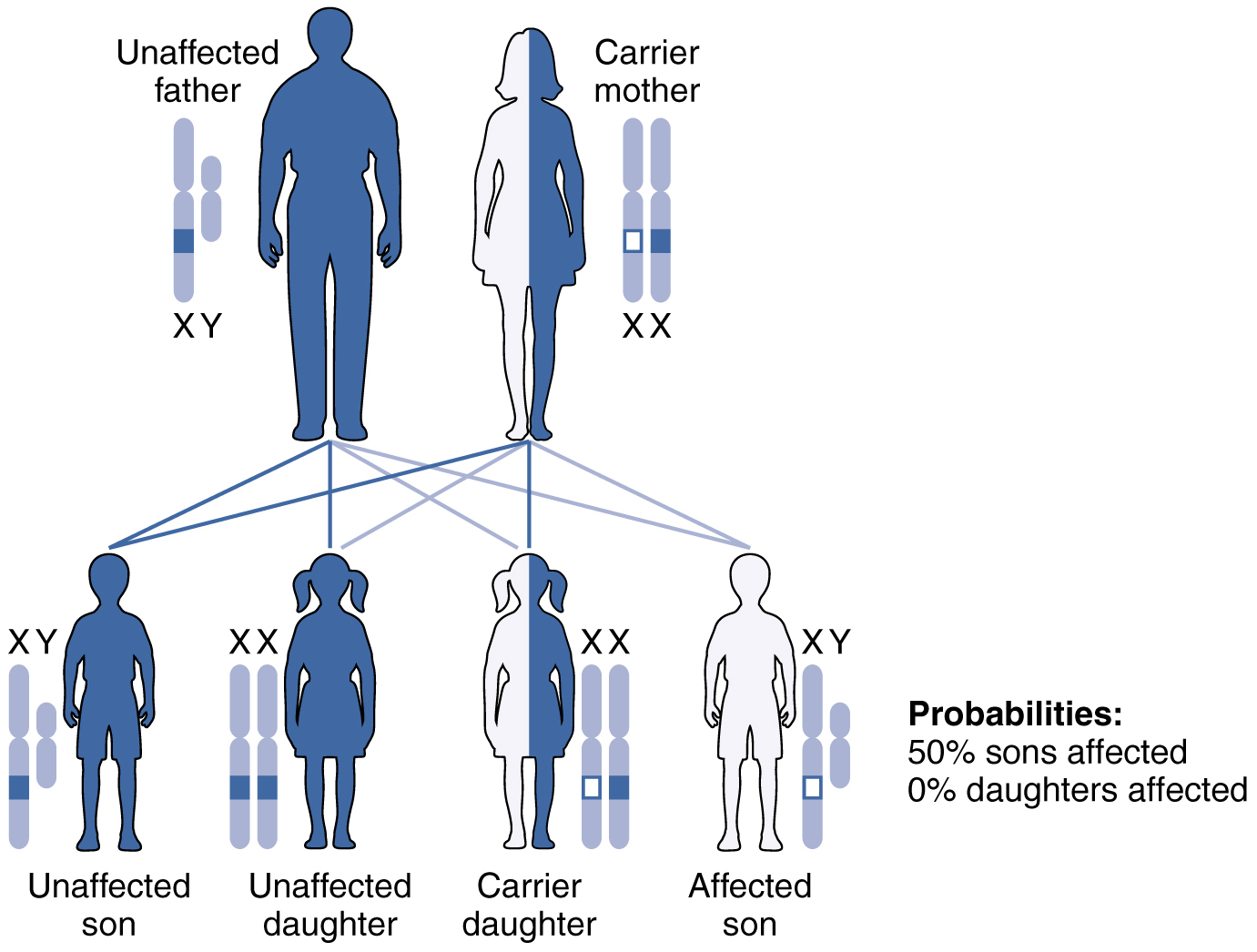
X-linked recessive inheritance is much more common because females can be carriers of the disease yet still have a normal phenotype. Diseases transmitted by X-linked recessive inheritance include color blindness, the blood-clotting disorder hemophilia, and some forms of muscular dystrophy. For an example of X-linked recessive inheritance, consider parents in which the mother is an unaffected carrier and the father is normal. None of the daughters would have the disease because they receive a normal gene from their father. However, they have a 50 percent chance of receiving the disease gene from their mother and becoming a carrier. In contrast, 50 percent of the sons would be affected (Figure 28.7.6).
With X-linked recessive diseases, males either have the disease or are genotypically normal—they cannot be carriers. Females, however, can be genotypically normal, a carrier who is phenotypically normal, or affected with the disease. A daughter can inherit the gene for an X-linked recessive illness when her mother is a carrier or affected, or her father is affected. The daughter will be affected by the disease only if she inherits an X-linked recessive gene from both parents. As you can imagine, X-linked recessive disorders affect many more males than females. For example, color blindness affects at least 1 in 20 males, but only about 1 in 400 females.
Other Inheritance Patterns: Incomplete Dominance, Codominance, and Lethal Alleles
Not all genetic disorders are inherited in a dominant–recessive pattern. In incomplete dominance, the offspring express a heterozygous phenotype that is intermediate between one parent’s homozygous dominant trait and the other parent’s homozygous recessive trait. An example of this can be seen in snapdragons when red-flowered plants and white-flowered plants are crossed to produce pink-flowered plants. In humans, incomplete dominance occurs with one of the genes for hair texture. When one parent passes a curly hair allele (the incompletely dominant allele) and the other parent passes a straight-hair allele, the effect on the offspring will be intermediate, resulting in hair that is wavy.
Codominance is characterized by the equal, distinct, and simultaneous expression of both parents’ different alleles. This pattern differs from the intermediate, blended features seen in incomplete dominance. A classic example of codominance in humans is ABO blood type. People are blood type A if they have an allele for an enzyme that facilitates the production of surface antigen A on their erythrocytes. This allele is designated IA. In the same manner, people are blood type B if they express an enzyme for the production of surface antigen B. People who have alleles for both enzymes (IA and IB) produce both surface antigens A and B. As a result, they are blood type AB. Because the effect of both alleles (or enzymes) is observed, we say that the IA and IB alleles are codominant. There is also a third allele that determines blood type. This allele (i) produces a nonfunctional enzyme. People who have two i alleles do not produce either A or B surface antigens: they have type O blood. If a person has IA and i alleles, the person will have blood type A. Notice that it does not make any difference whether a person has two IA alleles or one IA and one i allele. In both cases, the person is blood type A. Because IA masks i, we say that IA is dominant to i. Table 28.4 summarizes the expression of blood type.
| Expression of Blood Types (Table 28.4) | ||
|---|---|---|
| Blood type | Genotype | Pattern of inheritance |
| A | IAIA or IAi | IAis dominant to i |
| B | IBIB orIBi | IB is dominant to i |
| AB | IAIB | IA is co-dominant to IB |
| O | ii | Two recessive alleles |
Certain combinations of alleles can be lethal, meaning they prevent the individual from developing in utero, or cause a shortened life span. In recessive lethal inheritance patterns, a child who is born to two heterozygous (carrier) parents and who inherited the faulty allele from both would not survive. An example of this is Tay–Sachs, a fatal disorder of the nervous system. In this disorder, parents with one copy of the allele for the disorder are carriers. If they both transmit their abnormal allele, their offspring will develop the disease and will die in childhood, usually before age 5.
Dominant lethal inheritance patterns are much more rare because neither heterozygotes nor homozygotes survive. Of course, dominant lethal alleles that arise naturally through mutation and cause miscarriages or stillbirths are never transmitted to subsequent generations. However, some dominant lethal alleles, such as the allele for Huntington’s disease, cause a shortened life span but may not be identified until after the person reaches reproductive age and has children. Huntington’s disease causes irreversible nerve cell degeneration and death in 100 percent of affected individuals, but it may not be expressed until the individual reaches middle age. In this way, dominant lethal alleles can be maintained in the human population. Individuals with a family history of Huntington’s disease are typically offered genetic counseling, which can help them decide whether or not they wish to be tested for the faulty gene.
Mutations
A mutation is a change in the sequence of DNA nucleotides that may or may not affect a person’s phenotype. Mutations can arise spontaneously from errors during DNA replication, or they can result from environmental insults such as radiation, certain viruses, or exposure to tobacco smoke or other toxic chemicals. Because genes encode for the assembly of proteins, a mutation in the nucleotide sequence of a gene can change amino acid sequence and, consequently, a protein’s structure and function. Spontaneous mutations occurring during meiosis are thought to account for many spontaneous abortions (miscarriages).
Chromosomal Disorders
Sometimes a genetic disease is not caused by a mutation in a gene, but by the presence of an incorrect number of chromosomes. For example, Down syndrome is caused by having three copies of chromosome 21. This is known as trisomy 21. The most common cause of trisomy 21 is chromosomal nondisjunction during meiosis. The frequency of nondisjunction events appears to increase with age, so the frequency of bearing a child with Down syndrome increases in women over 36. The age of the father matters less because nondisjunction is much less likely to occur in a sperm than in an egg.
Whereas Down syndrome is caused by having three copies of a chromosome, Turner syndrome is caused by having just one copy of the X chromosome. This is known as monosomy. The affected child is always female. Women with Turner syndrome are sterile because their sexual organs do not mature.
Career Connections
Genetic Counselor
Given the intricate orchestration of gene expression, cell migration, and cell differentiation during prenatal development, it is amazing that the vast majority of newborns are healthy and free of major birth defects. When a woman over 35 is pregnant or intends to become pregnant, or her partner is over 55, or if there is a family history of a genetic disorder, she and her partner may want to speak to a genetic counselor to discuss the likelihood that their child may be affected by a genetic or chromosomal disorder. A genetic counselor can interpret a couple’s family history and estimate the risks to their future offspring.
For many genetic diseases, a DNA test can determine whether a person is a carrier. For instance, carrier status for Fragile X, an X-linked disorder associated with mental retardation, or for cystic fibrosis can be determined with a simple blood draw to obtain DNA for testing. A genetic counselor can educate a couple about the implications of such a test and help them decide whether to undergo testing. For chromosomal disorders, the available testing options include a blood test, amniocentesis (in which amniotic fluid is tested), and chorionic villus sampling (in which tissue from the placenta is tested). Each of these has advantages and drawbacks. A genetic counselor can also help a couple cope with the news that either one or both partners is a carrier of a genetic illness, or that their unborn child has been diagnosed with a chromosomal disorder or other birth defect.
To become a genetic counselor, one needs to complete a 4-year undergraduate program and then obtain a Master of Science in Genetic Counseling from an accredited university. Board certification is attained after passing examinations by the American Board of Genetic Counseling. Genetic counselors are essential professionals in many branches of medicine, but there is a particular demand for preconception and prenatal genetic counselors.
External Website

Visit the National Society of Genetic Counselors website for more information about genetic counselors.
External Website

Visit the American Board of Genetic Counselors, Inc., website for more information about genetic counselors.
Chapter Review
There are two aspects to a person’s genetic makeup. Their genotype refers to the genetic makeup of the chromosomes found in all their cells and the alleles that are passed down from their parents. Their phenotype is the expression of that genotype, based on the interaction of the paired alleles, as well as how environmental conditions affect that expression.
Working with pea plants, Mendel discovered that the factors that account for different traits in parents are discretely transmitted to offspring in pairs, one from each parent. He articulated the principles of random segregation and independent assortment to account for the inheritance patterns he observed. Mendel’s factors are genes, with differing variants being referred to as alleles and those alleles being dominant or recessive in expression. Each parent passes one allele for every gene on to offspring, and offspring are equally likely to inherit any combination of allele pairs. When Mendel crossed heterozygous individuals, he repeatedly found a 3:1 dominant–recessive ratio. He correctly postulated that the expression of the recessive trait was masked in heterozygotes but would resurface in their offspring in a predictable manner.
Human genetics focuses on identifying different alleles and understanding how they express themselves. Medical researchers are especially interested in the identification of inheritance patterns for genetic disorders, which provides the means to estimate the risk that a given couple’s offspring will inherit a genetic disease or disorder. Patterns of inheritance in humans include autosomal dominance and recessiveness, X-linked dominance and recessiveness, incomplete dominance, codominance, and lethality. A change in the nucleotide sequence of DNA, which may or may not manifest in a phenotype, is called a mutation.
Review Questions
Critical Thinking Questions
1. Explain why it was essential that Mendel perform his crosses using a large sample size?
2. How can a female carrier of an X-linked recessive disorder have a daughter who is affected?
Glossary
- allele
- alternative forms of a gene that occupy a specific locus on a specific gene
- autosomal chromosome
- in humans, the 22 pairs of chromosomes that are not the sex chromosomes (XX or XY)
- autosomal dominant
- pattern of dominant inheritance that corresponds to a gene on one of the 22 autosomal chromosomes
- autosomal recessive
- pattern of recessive inheritance that corresponds to a gene on one of the 22 autosomal chromosomes
- carrier
- heterozygous individual who does not display symptoms of a recessive genetic disorder but can transmit the disorder to his or her offspring
- codominance
- pattern of inheritance that corresponds to the equal, distinct, and simultaneous expression of two different alleles
- dominant
- describes a trait that is expressed both in homozygous and heterozygous form
- dominant lethal
- inheritance pattern in which individuals with one or two copies of a lethal allele do not survive in utero or have a shortened life span
- genotype
- complete genetic makeup of an individual
- heterozygous
- having two different alleles for a given gene
- homozygous
- having two identical alleles for a given gene
- incomplete dominance
- pattern of inheritance in which a heterozygous genotype expresses a phenotype intermediate between dominant and recessive phenotypes
- karyotype
- systematic arrangement of images of chromosomes into homologous pairs
- mutation
- change in the nucleotide sequence of DNA
- phenotype
- physical or biochemical manifestation of the genotype; expression of the alleles
- Punnett square
- grid used to display all possible combinations of alleles transmitted by parents to offspring and predict the mathematical probability of offspring inheriting a given genotype
- recessive
- describes a trait that is only expressed in homozygous form and is masked in heterozygous form
- recessive lethal
- inheritance pattern in which individuals with two copies of a lethal allele do not survive in utero or have a shortened life span
- sex chromosomes
- pair of chromosomes involved in sex determination; in males, the XY chromosomes; in females, the XX chromosomes
- trait
- variation of an expressed characteristic
- X-linked
- pattern of inheritance in which an allele is carried on the X chromosome of the 23rd pair
- X-linked dominant
- pattern of dominant inheritance that corresponds to a gene on the X chromosome of the 23rd pair
- X-linked recessive
- pattern of recessive inheritance that corresponds to a gene on the X chromosome of the 23rd pair
Solutions
Answers for Critical Thinking Questions
- By using large sample sizes, Mendel minimized the effect of random variability resulting from chance. This allowed him to identify true ratios corresponding to dominant–recessive inheritance.
- The only way an affected daughter could be born is if the female carrier mated with a male who was affected. In this case, 50 percent of the daughters would be affected. Alternatively, but exceedingly unlikely, the daughter could become affected by a spontaneous mutation.
This work, Anatomy & Physiology, is adapted from Anatomy & Physiology by OpenStax, licensed under CC BY. This edition, with revised content and artwork, is licensed under CC BY-SA except where otherwise noted.
Images, from Anatomy & Physiology by OpenStax, are licensed under CC BY except where otherwise noted.
Access the original for free at https://openstax.org/books/anatomy-and-physiology/pages/1-introduction.
