25.7 Physiology of Urine Formation: Regulation of Fluid Volume and Composition
Learning Objectives
By the end of this section, you will be able to:
- Explain the mechanism of action of diuretics
- Explain why the differential permeability or impermeability of specific sections of the nephron tubules is necessary for urine formation
- Describe how sodium, potassium, chloride, calcium, phosphate, hydrogen ion, bicarbonate, ph, and nitrogenous wastes are regulated.
The major hormones influencing total body water are ADH, aldosterone, and ANH. Circumstances that lead to fluid depletion in the body include blood loss and dehydration. Homeostasis requires that volume and osmolarity be preserved. Blood volume is important in maintaining sufficient blood pressure, and there are nonrenal mechanisms involved in its preservation, including vasoconstriction, which can act within seconds of a drop in pressure. Thirst mechanisms are also activated to promote the consumption of water lost through respiration, evaporation, or urination. Hormonal mechanisms are activated to recover volume while maintaining a normal osmotic environment. These mechanisms act principally on the kidney.
Volume-sensing Mechanisms
The body cannot directly measure blood volume, but blood pressure can be measured. Blood pressure often reflects blood volume and is measured by baroreceptors in the aorta and carotid sinuses. When blood pressure increases, baroreceptors send more frequent action potentials to the central nervous system, leading to widespread vasodilation. Included in this vasodilation are the afferent arterioles supplying the glomerulus, resulting in increased GFR, and water loss by the kidneys. If pressure decreases, fewer action potentials travel to the central nervous system, resulting in more sympathetic stimulation-producing vasoconstriction, which will result in decreased filtration and GFR, and water loss.
Decreased blood pressure is also sensed by the granular cells in the afferent arteriole of the JGA. In response, the enzyme renin is released. You saw earlier in the chapter that renin activity leads to an almost immediate rise in blood pressure as activated angiotensin II produces vasoconstriction. The rise in pressure is sustained by the aldosterone effects initiated by angiotensin II; this includes an increase in Na+ retention and water volume. As an aside, late in the menstrual cycle, progesterone has a modest influence on water retention. Due to its structural similarity to aldosterone, progesterone binds to the aldosterone receptor in the collecting duct of the kidney, causing the same, albeit weaker, effect on Na+ and water retention.
Cardiomyocytes of the atria also respond to greater stretch (as blood pressure rises) by secreting ANH. ANH opposes the action of aldosterone by inhibiting the recovery of Na+ by the DCT and collecting ducts. More Na+ is lost, and as water follows, total blood volume and pressure decline. In low-pressure states, ANH does not seem to have much effect.
ADH is also called vasopressin. Early researchers found that in cases of unusually high secretion of ADH, the hormone caused vasoconstriction (vasopressor activity, hence the name). Only later were its antidiuretic properties identified. Synthetic ADH is still used occasionally to stem life-threatening esophagus bleeding in alcoholics.
When blood volume drops 5–10 percent, causing a decrease in blood pressure, there is a rapid and significant increase in ADH release from the posterior pituitary. Immediate vasoconstriction to increase blood pressure is the result. ADH also causes activation of aquaporin channels in the collecting ducts to affect the recovery of water to help restore vascular volume.
Diuretics and Fluid Volume
A diuretic is a compound that increases urine volume. Three familiar drinks contain diuretic compounds: coffee, tea, and alcohol. The caffeine in coffee and tea works by promoting vasodilation in the nephron, which increases GFR. Alcohol increases GFR by inhibiting ADH release from the posterior pituitary, resulting in less water recovery by the collecting duct. In cases of high blood pressure, diuretics may be prescribed to reduce blood volume and, thereby, reduce blood pressure. The most frequently prescribed anti-hypertensive diuretic is hydrochlorothiazide. It inhibits the Na+/ Cl– symporter in the DCT and collecting duct. The result is a loss of Na+ with water following passively by osmosis.
Osmotic diuretics promote water loss by osmosis. An example is the indigestible sugar mannitol, which is most often administered to reduce brain swelling after head injury. However, it is not the only sugar that can produce a diuretic effect. In cases of poorly controlled diabetes mellitus, glucose levels exceed the capacity of the tubular glucose symporters, resulting in glucose in the urine. The unrecovered glucose becomes a powerful osmotic diuretic. Classically, in the days before glucose could be detected in the blood and urine, clinicians identified diabetes mellitus by the three Ps: polyuria (diuresis), polydipsia (increased thirst), and polyphagia (increased hunger).
Regulation of Extracellular Na+
Sodium has a very strong osmotic effect and attracts water. It plays a larger role in the osmolarity of the plasma than any other circulating component of the blood. If there is too much Na+ present, either due to poor control or excess dietary consumption, a series of metabolic problems ensue. There is an increase in total volume of water, which leads to hypertension (high blood pressure). Over a long period, this increases the risk of serious complications such as heart attacks, strokes, and aneurysms. It can also contribute to system-wide edema (swelling).
Mechanisms for regulating Na+ concentration include the renin–angiotensin–aldosterone system and ADH (see Chapter 25 Figure Figure 25.4.2). Aldosterone stimulates the uptake of Na+ on the apical cell membrane of cells in the DCT and collecting ducts, whereas ADH helps to regulate Na+ concentration indirectly by regulating the reabsorption of water.
Regulation of Extracellular K+
Potassium is present in a 30-fold greater concentration inside the cell than outside the cell. A generalization can be made that K+ and Na+ concentrations will move in opposite directions. When more Na+ is reabsorbed, more K+ is secreted; when less Na+ is reabsorbed (leading to excretion by the kidney), more K+ is retained. When aldosterone causes a recovery of Na+ in the nephron, a negative electrical gradient is created that promotes the secretion of K+ and Cl– into the lumen.
Regulation of Cl–
Chloride is important in acid–base balance in the extracellular space and has other functions, such as in the stomach, where it combines with hydrogen ions in the stomach lumen to form hydrochloric acid, aiding digestion. Its close association with Na+ in the extracellular environment makes it the dominant anion of this compartment, and its regulation closely mirrors that of Na+.
Regulation of Ca++ and Phosphate
The parathyroid glands monitor and respond to circulating levels of Ca++ in the blood. When levels drop too low, parathyroid hormone (PTH) is released to stimulate the DCT to reabsorb Ca++ from the forming urine. When levels are adequate or high, less PTH is released and more Ca++ remains in the forming urine to be lost. Phosphate levels move in the opposite direction. When Ca++ levels are low, PTH inhibits reabsorption of phosphate (PO3–) causing its loss in the urine which decreases the phosphate in the blood. The retention of phosphate would result in the formation of calcium phosphate in the plasma, reducing circulating Ca++ levels. By ridding the blood of phosphate, blood Ca++ levels rise. PTH also stimulates the renal conversion of calcidiol into calcitriol, the active form of vitamin D. Calcitriol then stimulates the intestines to absorb more Ca++ from the diet.
Regulation of H+, Bicarbonate, and pH
The acid–base homeostasis of the body is a function of chemical buffers and physiologic buffering provided by the lungs and kidneys. Buffers, especially proteins, HCO3−, and ammonia have a very large capacity to absorb or release H+ as needed to resist a change in pH. They can act within fractions of a second. The lungs can rid the body of excess acid very rapidly (seconds to minutes) through the conversion of HCO3– into CO2, which is then exhaled. It is rapid but has limited capacity in the face of a significant acid challenge. The kidneys can rid the body of both acid and base. The renal capacity is large but slow (minutes to hours). The cells of the PCT actively secrete H+ into the forming urine as Na+ is reabsorbed. The body rids itself of excess H+ and raises blood pH. In the collecting ducts, the apical surfaces of intercalated cells have proton pumps that actively secrete H+ into the luminal, forming urine to remove it from the body.
As hydrogen ions are pumped into the forming urine, it is buffered by bicarbonate (HCO3–), H2PO4– (dihydrogen phosphate ion), or ammonia (forming NH4+, ammonium ion). Urine pH typically varies in a normal range from 4.5 to 8.0.
Regulation of Nitrogen Wastes
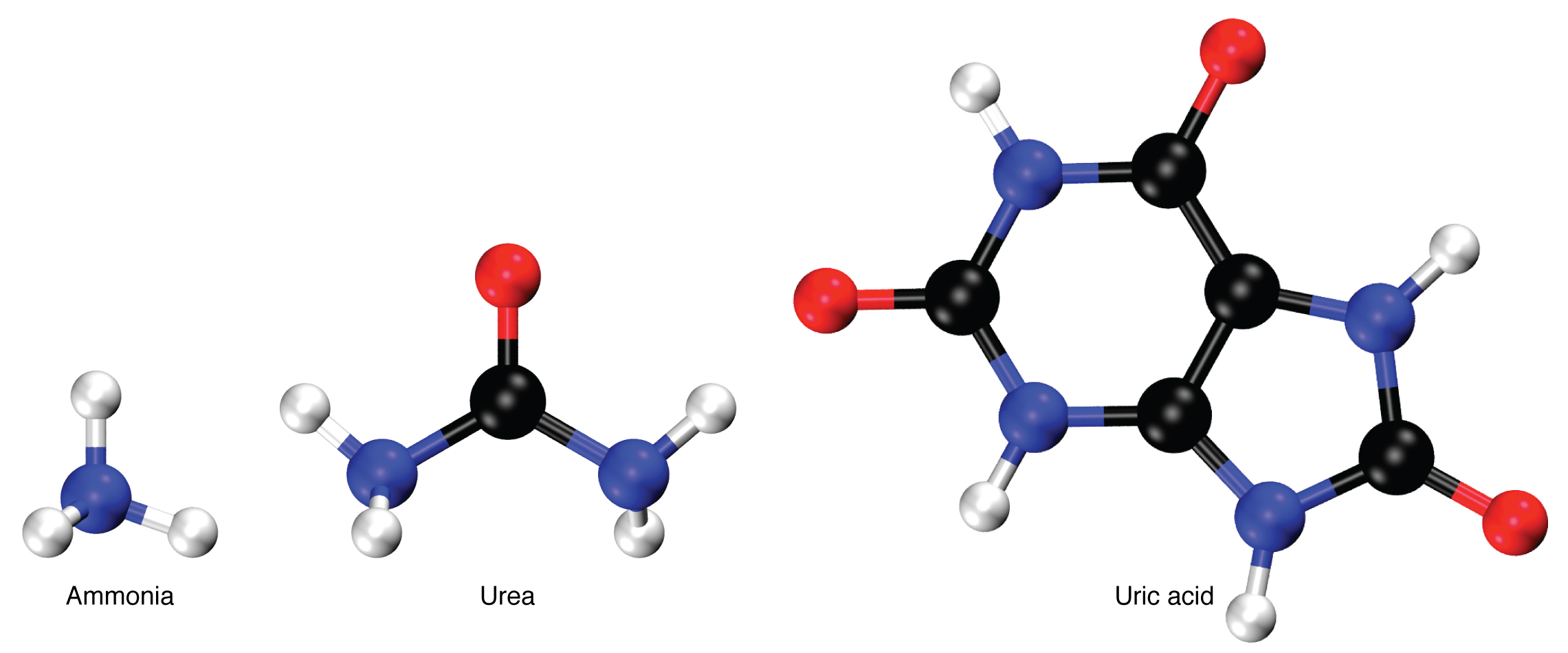
Nitrogen wastes are produced by the breakdown of proteins during normal metabolism. Proteins are broken down into amino acids, which in turn are deaminated by having their nitrogen groups removed. Deamination converts the amino (NH2) groups into ammonia (NH3), ammonium ion (NH4+), urea, or uric acid (Figure 25.7.1). Ammonia is extremely toxic, so most of it is very rapidly converted into urea in the liver. Human urinary wastes typically contain primarily urea with small amounts of ammonium and very little uric acid.
Elimination of Drugs and Hormones
Water-soluble drugs may be excreted in the urine and are influenced by one or all of the following processes: glomerular filtration, tubular secretion, or tubular reabsorption. Drugs that are structurally small can be filtered by the glomerulus with the filtrate. Large drug molecules such as heparin or those that are bound to plasma proteins cannot be filtered and are not readily eliminated. Some drugs can be eliminated by carrier proteins that enable secretion of the drug into the tubule lumen. There are specific carriers that eliminate basic (such as dopamine or histamine) or acidic drugs (such as penicillin or indomethacin). As is the case with other substances, drugs may be both filtered and reabsorbed passively along a concentration gradient.
Chapter Review
The major hormones regulating body fluids are ADH, aldosterone and ANH. Progesterone is similar in structure to aldosterone and can bind to and weakly stimulate aldosterone receptors, providing a similar but diminished response. Blood pressure is a reflection of blood volume and is monitored by baroreceptors in the aortic arch and carotid sinuses. When blood pressure increases, more action potentials are sent to the central nervous system, resulting in greater vasodilation, greater GFR, and more water lost in the urine. ANH is released by the cardiomyocytes when blood pressure increases, causing Na+ and water loss. ADH at high levels causes vasoconstriction in addition to its action on the collecting ducts to recover more water. Diuretics increase urine volume. Mechanisms for controlling Na+ concentration in the blood include the renin–angiotensin–aldosterone system and ADH. When Na+ is retained, K+ is excreted; when Na+ is lost, K+ is retained. When circulating Ca++ decreases, PTH stimulates the reabsorption of Ca++ and inhibits reabsorption of HPO42−. pH is regulated through buffers, expiration of CO2, and excretion of acid or base by the kidneys. The breakdown of amino acids produces ammonia. Most ammonia is converted into less-toxic urea in the liver and excreted in the urine. Regulation of drugs is by glomerular filtration, tubular secretion, and tubular reabsorption.
Review Questions
Critical Thinking Questions
1. Why is ADH also called vasopressin?
2. How can glucose be a diuretic?
Glossary
- diuretic
- compound that increases urine output, leading to decreased water conservation
Solutions
Answers for Critical Thinking Questions
- When first discovered, it was named for its known activity—vasoconstriction.
- In cases of diabetes mellitus, there is more glucose present than the kidney can recover and the excess glucose is lost in the urine. It possesses osmotic character so that it attracts water to the forming urine.
This work, Anatomy & Physiology, is adapted from Anatomy & Physiology by OpenStax, licensed under CC BY. This edition, with revised content and artwork, is licensed under CC BY-SA except where otherwise noted.
Images, from Anatomy & Physiology by OpenStax, are licensed under CC BY except where otherwise noted.
Access the original for free at https://openstax.org/books/anatomy-and-physiology/pages/1-introduction.
