Pinyon-Juniper Woodland Ecosystems
Pinyon-juniper (PJ) systems create a mosaic with sagebrush, pine, and montane ecosystems covering millions of acres. They are not well-studied systems, and they are often seen as barren “wasteland.” However, they most certainly are not: they are a dominant ecosystem in some of the West’s more beautiful landscapes and recreation areas. PJ systems are woodlands that have at least one Pinus species mixed with juniper (Juniperus spp.). Due to temperature tolerances, pinyon is generally not found beyond the reaches of northern Utah and southern portions of Wyoming and Idaho. PJ systems generally sit at mid-elevations on the landscape, with shrubland and grasslands at lower elevations and ponderosa pine, and montane PJ systems at higher elevations. Like most ecosystems in the West, PJ systems are stressed by a history of overgrazing, invasive species, and heavy recreational use. PJ systems are also under pressure from intensive natural gas extraction in southern Colorado and New Mexico.
Section 1: Geographic Distribution
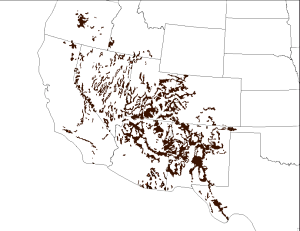
Pinyon-juniper woodlands are in regions subject to temperature extremes and limited precipitation throughout Nevada, Utah, Colorado, and New Mexico (figure 9.1). Geologic records indicate that around thirteen thousand years ago PJ systems moved north to areas of higher elevation as a result of climate conditions that were becoming drier and warmer.
Today PJ systems occur between 4,500 and 7,500 feet elevation and in areas with precipitation regimes that range from seven to twenty-five inches.
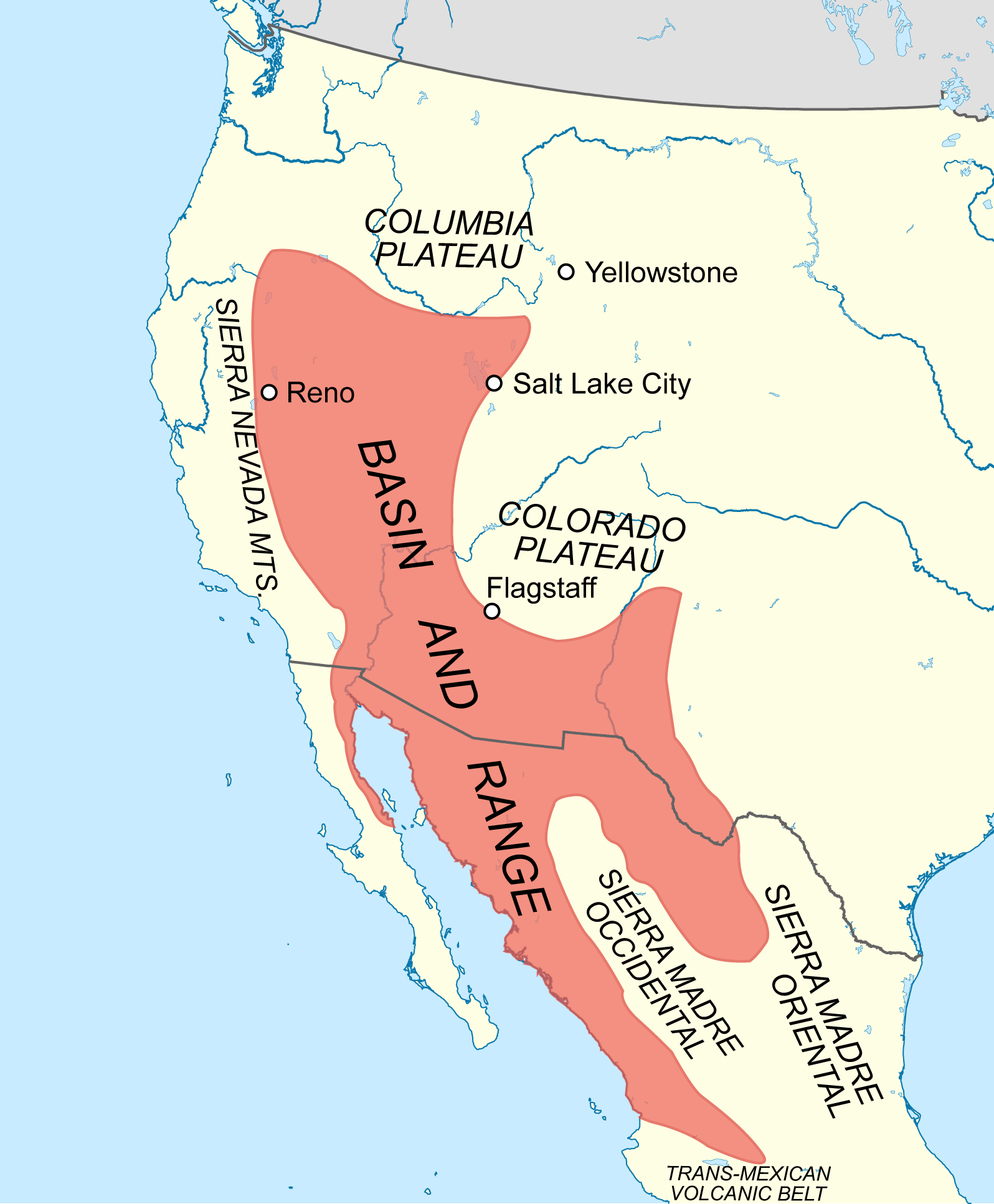
This chapter will also include systems in which PJ is present, but not dominant. These systems are mosaic gradients across elevations and include grasslands, shrublands, woodlands, and pine and alpine systems in transition zones between the Colorado Plateau province and the Basin and Range province (the area around the bold black line in Arizona and New Mexico in figure 9.2).
Section 2: Major Land Resource Areas (MLRAs) of Pinyon-Juniper Ecosystems
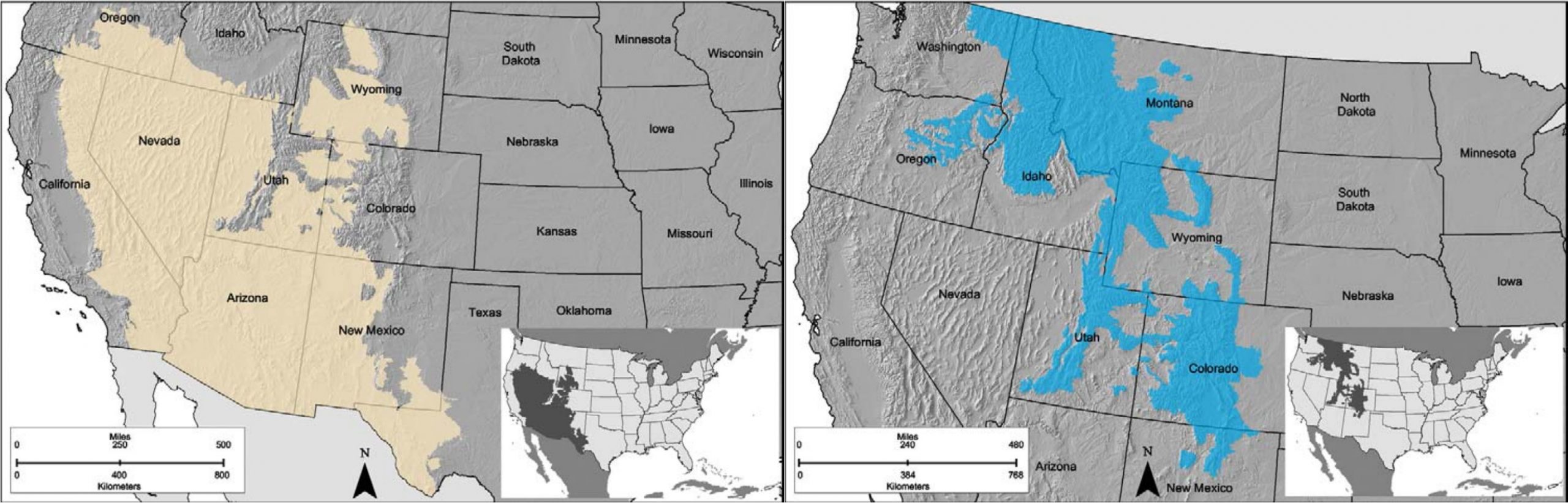
PJ systems are interspersed throughout sagebrush ecosystems in Land Resource Areas D and two regions of Land Resource Area E.
- Region D-35
- Region D-36
- Region D-38
- Region D-39
- Region E-49
- Region E-51
D-Region MLRAs
A general description of the D Land Resource Area is covered in chapter 8 on sagebrush systems.
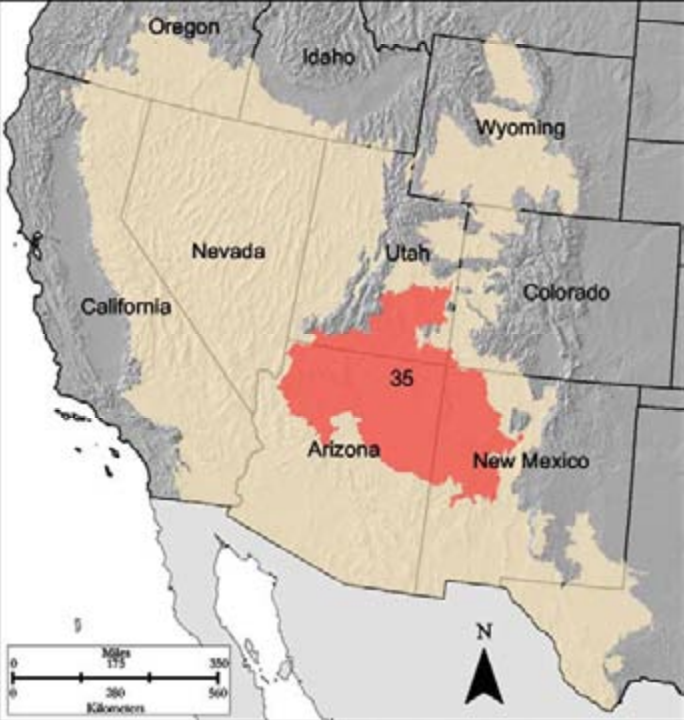
D-35 Colorado Plateau is located across the only location in the United States where a boundary of four states (UT, CO, AZ, NM) meet at a single point—Four Corners National Monument. This MLRA also encompasses multiple national parks and monuments, including the Grand Canyon, Canyonlands, Arches, and Grand Staircase–Escalante. Reservations of numerous Native American tribes are in this region, including Navajo, Hopi, Zuni, Hualapai, and Southern Utes. This region experiences a high volume of use as a tourist destination with many recreational and cultural opportunities. Vegetation across the region is a mosaic of sagebrush and PJ systems with understories of galleta (Hilaria rigida), blue grama (Bouteloua gracilis), black grama (Bouteloua eriopoda), and western wheatgrass. In lower elevation areas with alkaline soils vegetation includes alkali sacaton (Sporobolus airoides), Indian ricegrass (Oryzopsis hymenoides), needled grasses (Stipa spp.), four-wing saltbush (Atriplex canescens), and winterfat (Krascheninnikovia lanata). In areas with more saline soils, greasewood (Adenostoma fasciculatum) and shadescale (Atriplex confertifolia) are present among the sage, pinyon, juniper, and grasses.
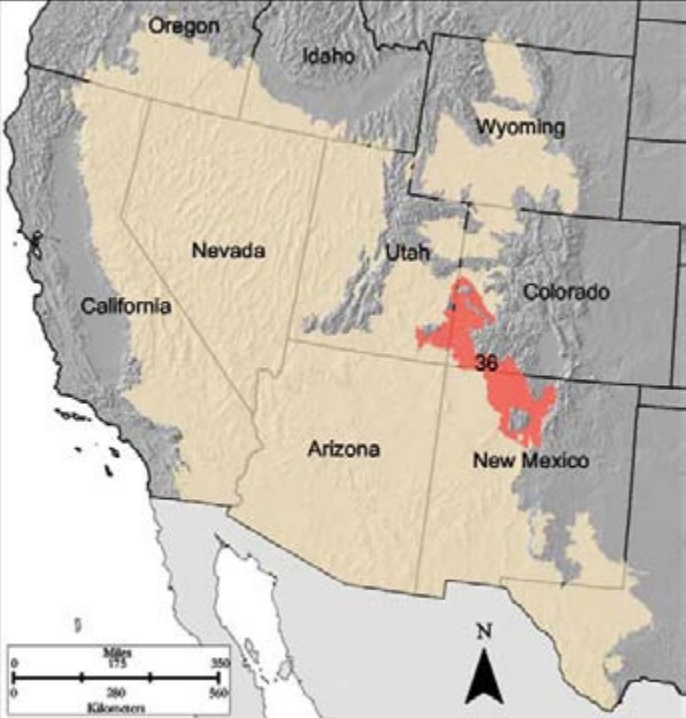

D-36 Southwestern Plateaus, Mesas, and Foothills is home to the author of this text as well as to the reservations of many tribes (Southern Ute, Jemez, Jicarilla Apache, Navajo, Santa Domingo, Zia, and others). This MLRA borders the Southern Rockies to the north. Vegetation is a mosaic across the elevation gradient (figure 9.6). Interspersed throughout the systems at lower elevations are Gambel Oak (Quercus gambelii), mountain muhly (Muhlenbergia montana), and snowberry (Symphoricarpos spp.).
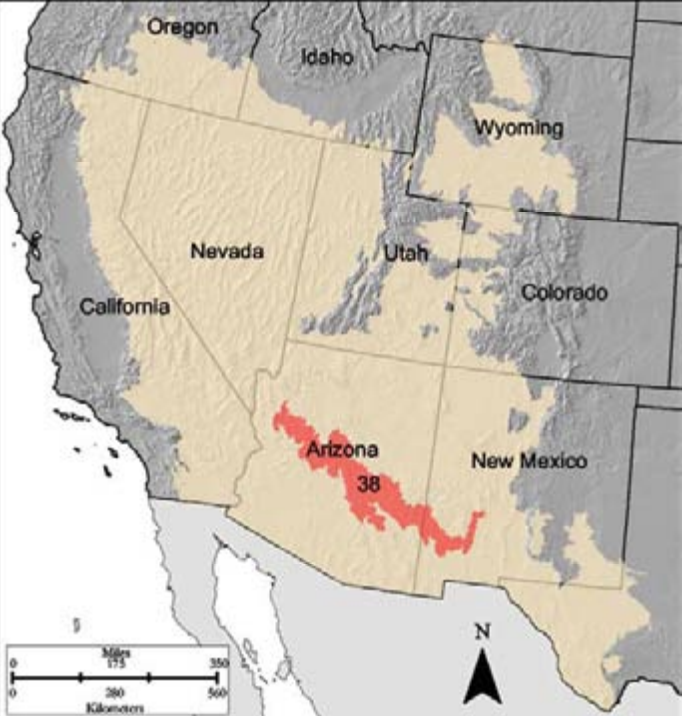
D-38 Mogollon Transition is a boundary diagonal strip of rugged low mountains interspersed with canyons, including the Salt River Canyon, across Central Arizona into the Gila National Forest area of southwest New Mexico between the Colorado Plateau and the southern portion of the Basin and Range province. Deep alluvium washed from surrounding mountains covers much of the lower elevation areas. A volcanic history with extensive basalt flows dominates the area. Higher elevations have granite peaks that are dated at over one billion years old.
Like MLRA D-36, D-38 has a mosaic gradient of shrubs to woodland to pine to fir systems. The pinyon in the PJ woodlands is largely Mexican pinyon pine (Pinus cembroides). Included in the PJ woodland system are Mexican blue oak (Quercus oblongifolia), New Mexico locust (Robinia neomexicana), buckbrush (Ceanothus cuneatus), and manzanita (Arctostaphylos spp.). Understory includes grama, needlegrasses, and jojoba, among a diversity of other grass and forb species. Given the MLRA’s location adjacent to the Sonoran Desert, diversity of plant, animal, bird, reptile, and insect species increases greatly over northern MLRAs of the D Land Resource Area.
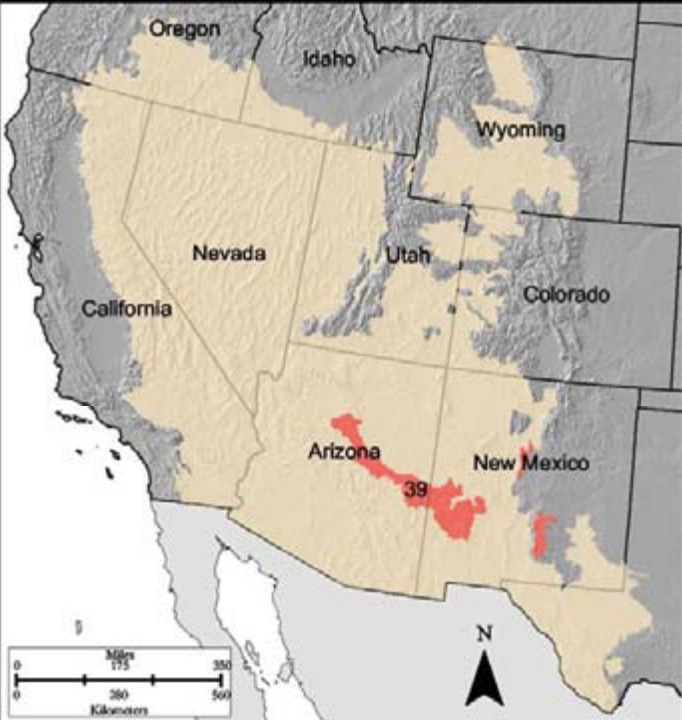
D-39 Arizona and New Mexico Mountains is just north of the Mollogon Rim and also stretches in a strip across Arizona into New Mexico, with a few small patches in Central New Mexico. The area includes areas of Cibola, Kaibab, and Gila National Forests as well as the two highest points in Arizona—Baldy Peak (11,403 feet) and Humphrey’s Peak (12,670 feet). Again, a vegetation gradient across elevations occurs in this region. At lower elevations of deep soils are grasslands in which gramas, needlegrasses, and June grasses (Koeleria spp.) dominate. Mid elevations are shrub-woodland systems comprised of oak, pine, and juniper species. North-facing slopes are dominated by PJ, while south-facing slopes contain PJ intermixed with oak and with understory of fescues, bluegrasses, and brome species. Woodland mixes with pine, which gives way to firs and spruces at higher elevations.
E-Region MLRAs
Much of Land Resource Region E is in the rugged areas and valleys of the Rocky Mountains spanning from the Canadian border down through northern New Mexico. Over half of the region is managed by the federal government for timber, grazing, and recreation. This text will focus on two MLRAs in the southern reaches of the region—E-49 and E-51.
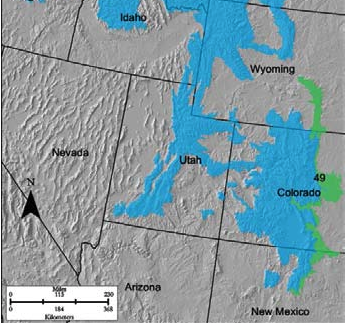
E-49 Southern Rocky Mountain Foothills could otherwise be known as the “Front Range” in Colorado. PJ systems in this MLRA are only found at lower elevations in the southern portion, extending down to Santa Fe, New Mexico, mixed with sagebrush, mountain mahogany (Cercocarpus spp.), grama and needled grasses, and western wheatgrass. This area is largely undeveloped at the moment, but the heavily traveled corridor of Interstate 25 runs through it. It is dotted with wind farms and grazing operations. The upper elevation forests are national forests and timber extraction operations.
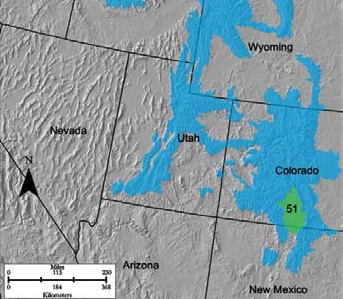
E-51 High Intermountain Valleys is a small patch that largely encompasses the San Luis Valley, with the moderately busy Highway 160 crossing through. This home to the Great Sand Dunes National Park and is bounded by the Sangre de Cristo Mountains and the San Juan Mountains. Agriculture, grazing, and recreation are dominant land uses. The vegetation community is a mosaic of shrub-grasslands and PJ woodlands. Shrubs include greasewood, rabbitbrush, four-wing saltbush, alkali sacaton, and big sagebrush. The understory is a mix of grama and needled grasses and western wheatgrass. Stream areas are lined with cottonwoods.
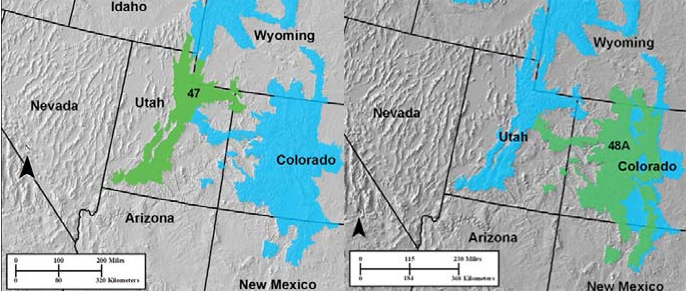
Two other MLRAs of note in this land resource area are E-47 Wasatch and Uinta Mountains and E-48A Southern Rocky Mountains (figure x). Both have patches of PJ systems in their respective elevation vegetation gradients from shrub-grasslands to woodlands to pine to alpine systems.
Alternative Classifications and Profiles of PJ Systems
In the few studies that have been conducted on PJ systems, most have focused on differences among the systems, and an informal classification scheme of savanna, woodland-shrubland, and persistent woodland has emerged. Table 9.1 differentiates among these three classifications across key variables.
| Type | Savanna | Wooded shrubland | Persistent woodland |
|---|---|---|---|
| Precipitation | Receives mainly summer precipitation; bimodal precipitation in some areas | Most often occurring (but not restricted to) areas of winter precipitation | Receives mainly winter precipitation |
| Soils | Deeper, fine-textured soils support a range of vegetation types | Wide variety of substrates | Shallow, often rocky soils do not support continuous vegetative cover |
| Terrain | Gentle terrain—valleys, basins, and foothills | Wide variety of topography—plains, valleys, and lower montane | Rugged upland sites and steep, rocky terrain |
| Stand structure | Open savanna-like stand structure, supports low density of trees and shrubs and dense herbaceous growth: grasses, forbs, and annuals | Areas of woodland expansion and contraction; shifts from herbaceous to shrub to tree dominance over time, in the absence of fire; often shrub dominated, with trees colonizing when growing conditions are favorable. | Multiaged stand structure. Range of tree densities and canopy cover, depending on site conditions |
| Historical fire regime | Frequent, lower-intensity surface fires maintain grasses and open stand structure; few barriers to fire spread | Infrequent, high-severity fires and patchy, mixed-severity fires | Infrequent high-severity fire. Significant barriers to fire spread: cliffs, canyons, exposed rock, topographic isolation |
| Impacts: Livestock and fire suppression | Increased density of younger trees, reduced ground cover, and higher-severity fires when they occur | Increased tree density, cover, and canopy closure; decreased shrub density; and larger higher-severity fires when they occur | Little impact to fire frequency and severity; understory species composition altered by livestock grazing |
Section 3: Pinyon Pine and Juniper Species
There are several species in each of the Pinus and Juniperus genera that comprise PJ woodlands and associated systems. There are eight species of true pinyon pine; four species of pinyon (no pine) species, and several bristlecone species. In the United States, there are three species of pinyon pines: single-leaf pinyon pine (Pinus monophylla), two-leaf or Colorado pinyon pine (Pinus edulis), and Mexican pinyon pine (Pinus cembroides). Overall, there are over fifty pinus species, six of which will be covered in this text.
Pinus Species
| Single-leaf pinyon pine
Pinus monophylla Green—subsp. monophylla Blue—subsp. californiarum Red—subsp. fallax  |
 |
| Two-leaf pinyon pine
(Colorado pinyon pine) Pinus edulis 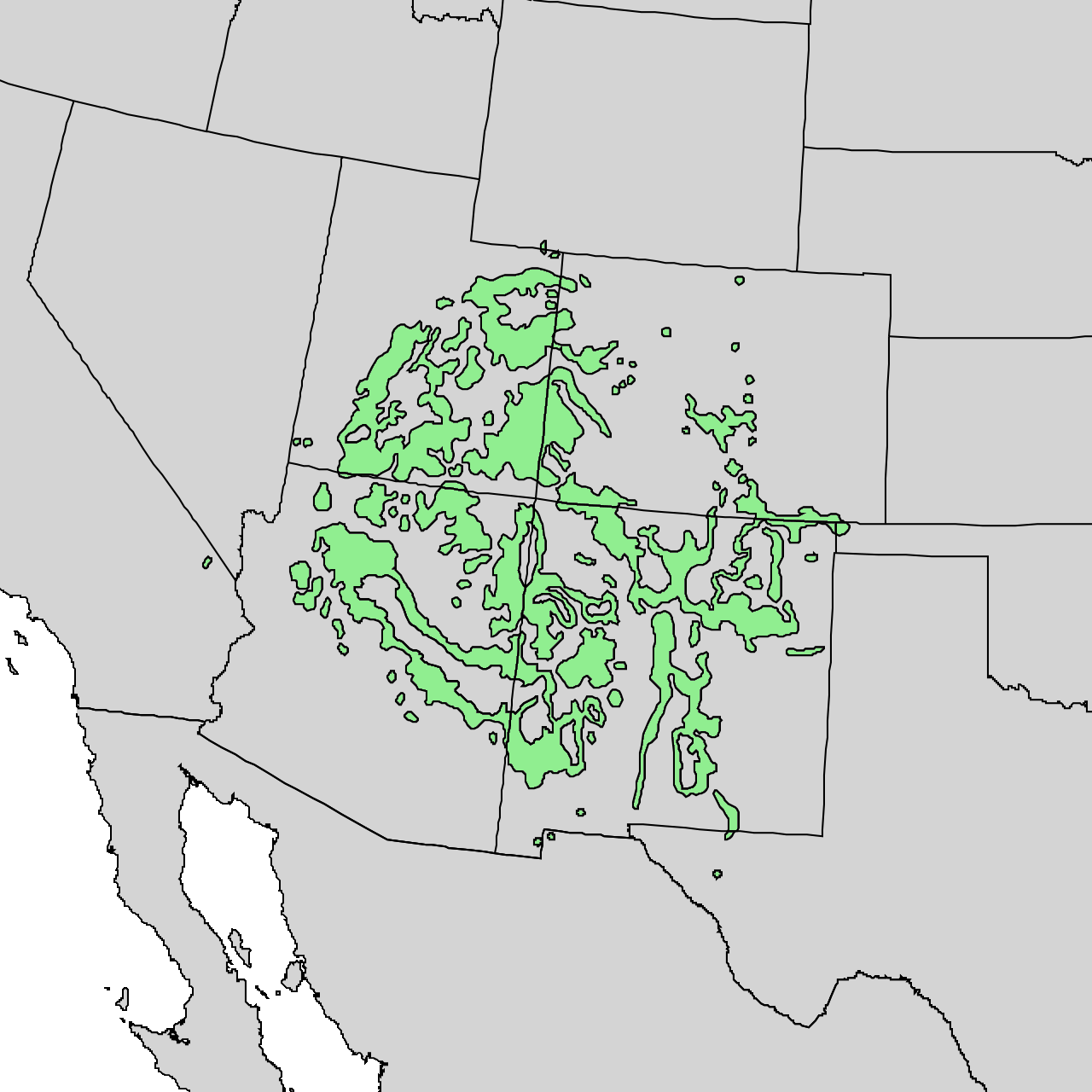 |
 |
| Mexican Pinyon Pine
Pinus cembroides 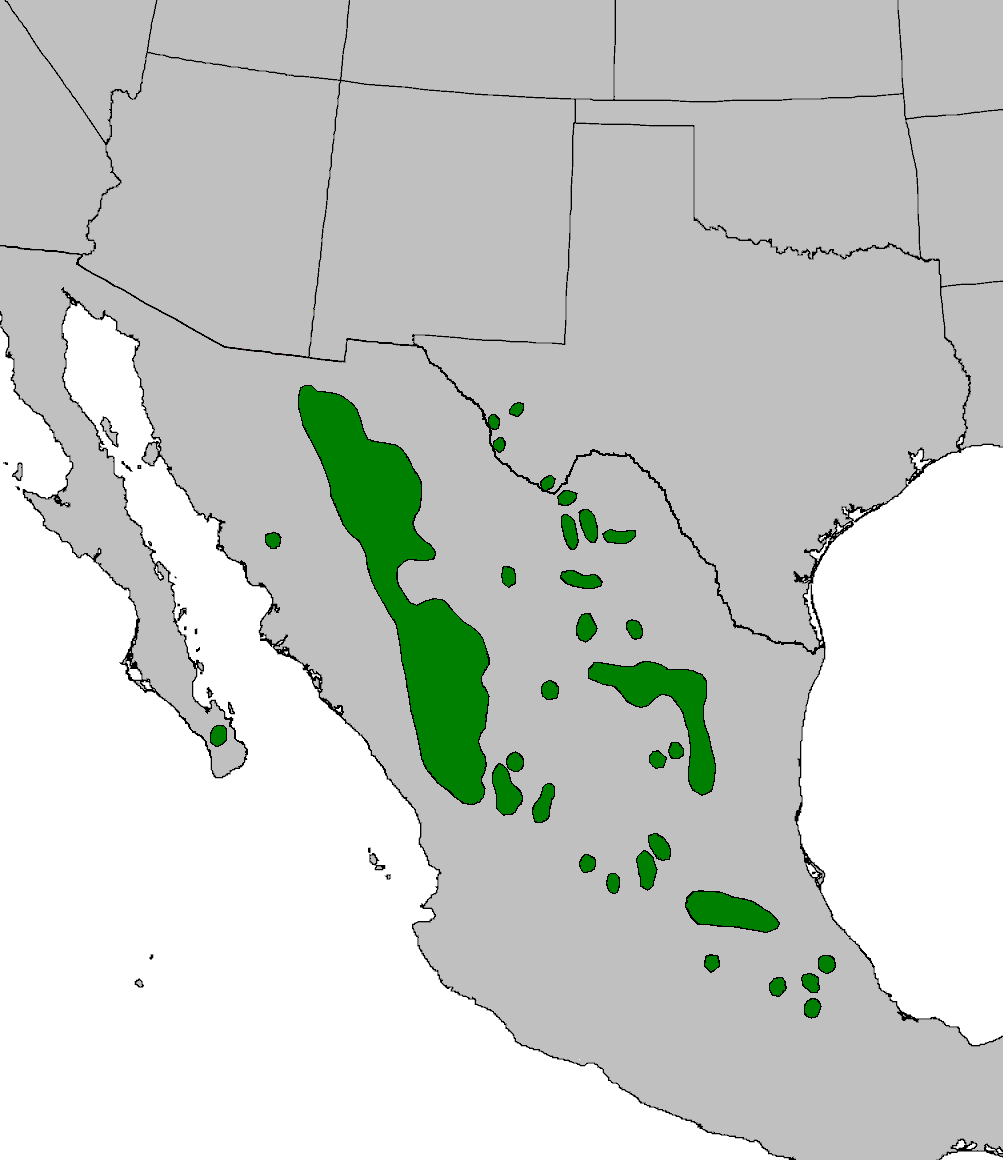 |
 |

Species in the Pinus genus are evergreen trees native to the western United States and ranging in mature height from twenty-five to seventy feet, depending on species; they have long waxy needles in various arrangements and produce small roundish cones. This species is known for its small, round, edible nut production, which has been a dietary staple for many Native American tribes throughout the Southwest. Many people burn pinyon pine as firewood, as it burns hot and has a rich, fragment aroma. Although species in the Pinus genus provide habitat to a number of bird and animal species, most notably is the pinyon jay (Gymnorhinus cyanocephalus; figure 9.18), which nests in pinyon trees and feeds on the nuts. These jays are a threatened species due to habitat fragmentation as land managers have historically attempted to convert extensive areas of PJ systems to grazing lands.
Juniperus Species
Several of the more than fifty species in the Juniperus genus are found in the PJ systems of the western United States. See figure 9.19.
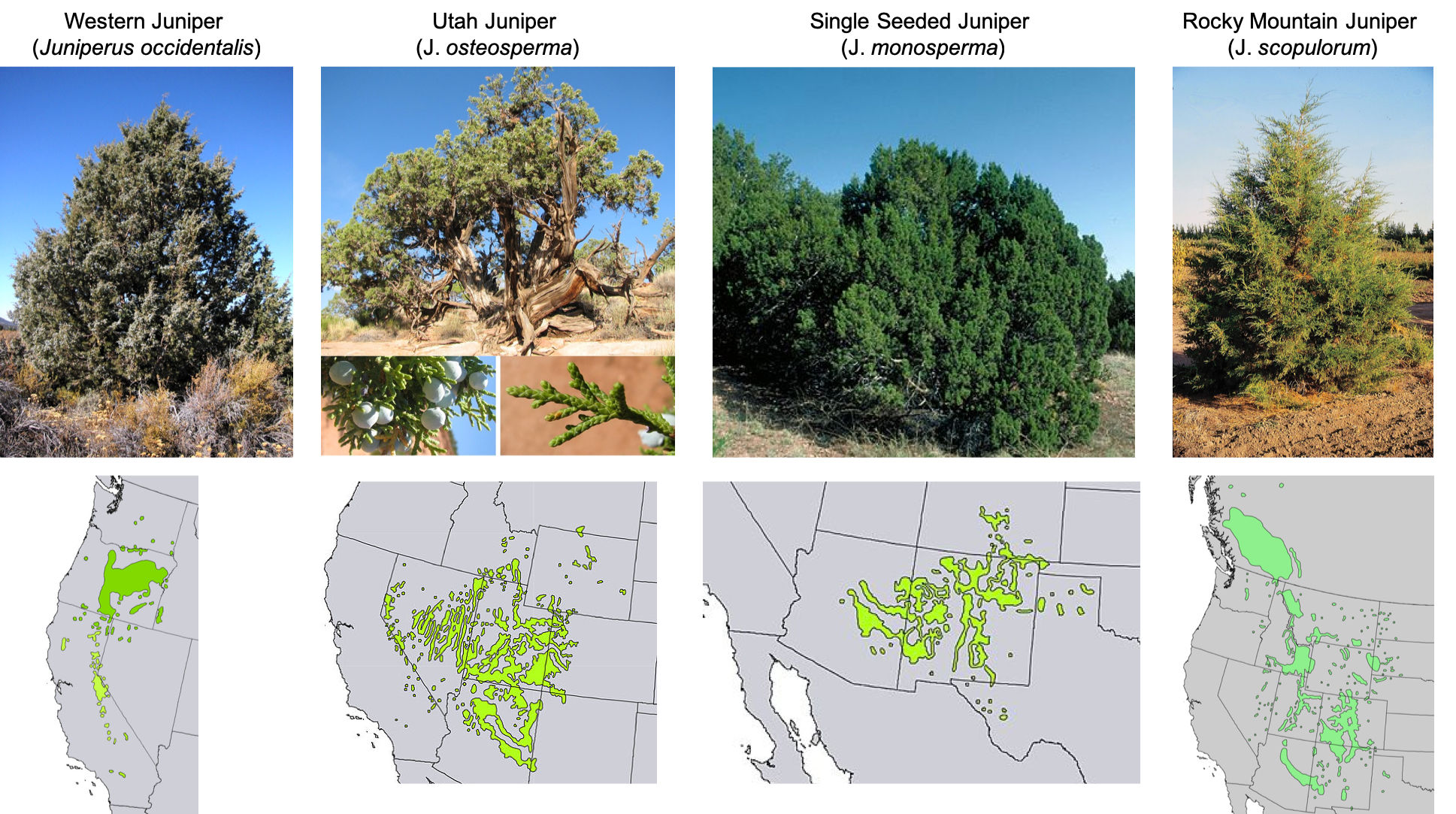
Species in the Juniperus genus range in mature height from thirty to seventy feet. Juniperus species are well known for slow growth habit, longevity, and distinctive twisting trunks (most species) of strong wood. The Juniperus genus includes some species that are monoecious (individual with both sexes) and others that are dioecious (single-sexed individuals). In some areas of the western United States, due to fire suppression and overgrazing, juniper is encroaching on grasslands and grass-shrub systems.
Drought Tolerance
Both pinyon and juniper tree species are adapted to tolerate a broad range of environmental conditions, enabling them to compete with a variety of plant communities and achieve wide distribution. Both mature pinyon and juniper trees have rooting systems that allow them to tolerate low moisture levels by extracting moisture through an extensive network of both lateral and taproots root systems (figure 9.20).
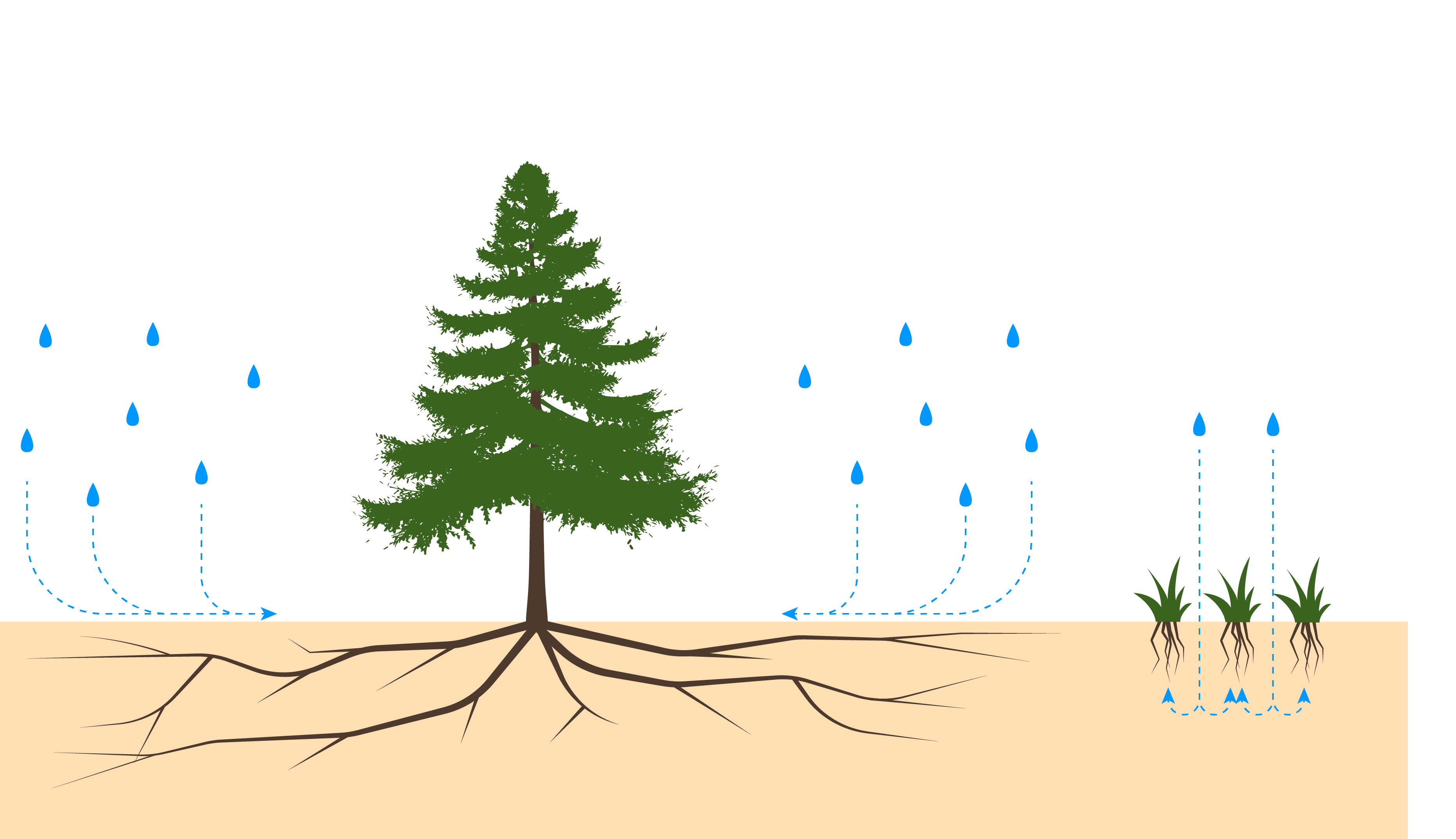
The deep taproots access moisture deep belowground that other plants, such as shrubs and grass, are unable to access. Their lateral roots systems, which are close to the soil surface, access moisture from recent precipitation events. The juniper tree species tends to remain at lower elevation levels and can tolerate drought more efficiently than can the pinyon tree species.
Section 4: Succession and Retrogression in Pinyon-Juniper Ecosystems
Expansions and contractions of pinyon-juniper habitat can be driven by factors, or combinations of factors, such as climate, insects, disease, and fire frequency. Climate and fire also influence pinyon pine and juniper succession, since certain conditions are better than others for seed germination and seedling establishment.
Climate and Drought
Moisture availability and temperature are important factors influencing the expansion and decline of pinyon-juniper woodlands. Surges of pinyon pine and juniper seedling recruitment and growth generally correspond with periods of wet climatic conditions, creating higher-density stands with greater fuel continuity.

Subsequent periods of drought create conditions conducive to large, stand-replacing fires. One of the most severe droughts of the twentieth century occurred between 1950 and 1956 in the southwestern United States and Mexico, causing massive die-off of many types of drought-adapted vegetation, including conifers, grasses, and cacti (Betancourt et al.1993). Two decades of higher than usual precipitation, followed by drought and combined with high winds have fueled six large fires in Mesa Verde National Park since 1996 (Floyd et al. 2000).
Periods of extreme drought intensify a number of other disturbance processes and ecological effects. Shaw et al. (2005) attribute a recent increase in pinyon mortality across portions of the Southwest to “a complex of drought, insects, and disease”—the cumulative effects of multiple mortality agents working in concert.
Endemic Insects and Disease
Pinyon pines affected by moisture stress are more susceptible to outbreaks of disease, parasitism by pinyon dwarf mistletoe (Arceuthobium divaricatum), and damage from endemic insects that feed on cones, needles, shoots, roots, and conductive tissue. Bark beetle and other insect outbreaks are closely tied to drought-induced moisture stress (Allen and Breshears 1998). Populations of pinyon tip moth (Dioryctria albovittella), pinyon cone moth (Eucosma bobana), pinyon Ips (Ips confusus), pinyon twig beetles (Pityophthorus spp. and Pityogenes spp.), pinyon needle scale (Matsucoccus acalyptus), and pinyon needle miner (Coleotechnites edulicola), normally present in pinyon-juniper woodlands at low levels, can increase dramatically during periods of drought, causing large-scale defoliation, reduced growth and reproduction, tree mortality, and a myriad of other effects. Juniper is also parasitized by its own species of mistletoe, the juniper mistletoe (Phoradendron juniperinum).
Fire
Wildfire has played an important role in structuring pinyon-juniper woodlands, with different fire regimes historically present in the three pinyon-juniper woodland types described in table 9.1 in section 2.3. Fires are described in terms of their severity, which is determined by the fire’s effect on vegetation and is usually defined in terms of the survival of dominant trees. Three major types of wildfire influence the range, age class, species composition, structure, and density of pinyon-juniper woodlands: (1) low-severity surface fires, (2) high-severity fires, and (3) mixed-severity fires. Low-severity surface fires are fueled primarily by dense herbaceous biomass, leaving most overstory trees alive. High-severity, stand-replacing fires can burn over the surface, or up in the tree crowns (crown fire), killing most trees and initiating stand replacement. Finally, mixed-severity fires are a combination of low-severity surface fires in some areas and high-severity fire in other areas, leaving a patchwork of surviving trees interspersed with patches of tree mortality where shrubs and herbaceous ground cover regenerate first.
Modeling historic fire regimes is difficult in PJ systems as neither pinyon nor juniper reliably forms fire scars as do ponderosa pine and other conifer species. Both pinyon pines and junipers, with thin bark and low crowns, are easily killed by wildfire. With the exception of young alligator juniper, pinyon and juniper species do not resprout after fire as do associated species of shrubs and oaks but take several decades to become reestablished by seed. Although dead juniper snags can be persistent on the landscape, they are difficult for researchers to age.
The introduction and expansion of livestock grazing throughout much of the Southwest removed the fine herbaceous fuels that create a continuous layer of combustible material over the surface of the landscape. Combined with fire detection and suppression efforts in the twentieth century, fire frequency has been reduced, but fires have tended to burn with greater intensity when they do occur. The frequency of large, severe fires has increased over the past twenty years in pinyon-juniper woodlands and other vegetation communities across the western United States (Romme et al. 2008).
Succession
Seed Production and Establishment
Pinyon pine reproduces only by seed, not vegetatively. Heavy crops of pinyon pine cones (called mast crops) vary by species and are produced every two to six years, followed by multiple years of low cone production (Betancourt et al. 1993). On average, mast crops in pinyon pines will occur in one to three out of every ten years (Gori and Bate 2007). The cycle of flowering, fruiting, and seed maturation takes three years, with seedlings becoming established four growing seasons after cone initiation (Betancourt et al. 1993). Pinyon seedlings are shade-tolerant and have the best chance of establishment and survival under the protected microclimates of “nurse plants.”
Juniper exhibits a similar seed-production cycle, where heavy (mast) crops of juniper berries/seed are produced every two to five years, interspersed with intervals of low seed production (Gori and Bate 2007). Young alligator juniper trees are able to sprout prolifically after the death of the main trunk, but the ability to resprout declines with age (Gottfried et al. 1995; Tirmenstein 1999). One-seed, redberry, Utah, Rocky Mountain, and mature alligator juniper must rely on seed production and dispersal for reestablishment. For both pinyon and juniper, levels of germination and establishment are highest when a mast seed crop is followed by a period of increased precipitation and the absence of fire (Gori and Bate 2007).
Herbivory and Seed Dispersal
Birds and small mammals, including chipmunks, raccoons, and squirrels feed on and cache juniper berries and pinyon nuts. Birds are considered to be the primary dispersal agents for juniper (Gottfried et al. 1995). Seeds that have passed through birds’ digestive tracts germinate faster than uneaten seeds (Johnsen 1962). Bird species responsible for dispersing pinyon seeds include scrub jays (Aphelocoma californica), pinyon jays (Gymnorhinus cyanocephalus), Steller’s jays (Cyanocitta stelleri), and Clark’s nutcrackers (Nucifraga columbiana). Seeds deposited or cached under the protective microclimate of nurse plants or other debris are more likely to germinate and become established than those deposited or cached in the open. One potential advantage of seed masting is that surplus seeds remain after seed predators have consumed a portion of that year’s crop.
Succession after Fire
Generally, after a stand-replacing fire, succession begins with the establishment of annual grasses and forbs for several years, followed by perennial grasses and forbs. Shrubs are the next cover type to become established, utilizing the increased humidity and buffering from the sun and wind provided by perennial vegetation. In the absence of fire, juniper and pinyon will become the climax species, but they may take several decades or longer to become established. As canopy cover increases, shrubs and herbaceous cover tend to decrease in density and abundance. Shrubs may remain a component of pinyon-juniper woodland indefinitely or may eventually be suppressed by the slow-growing overstory, which takes hundreds of years to reach the “open” old-growth stage.
After a low-severity surface fire, surviving shrubs may regenerate quickly by resprouting. Frequent low-severity fires can eliminate pinyon and juniper seedlings and maintain shrub-dominated plant communities.
State and Transition Model
In 2006, the Nature Conservancy undertook an initiative to understand and model historic, current, and potential vegetation changes over time throughout the Southwest under the impact of varying fire intensities and concomitant impacts of drought and insect infestation. The initiative looked at a spectrum of systems from semidesert grasslands to shrublands to woodlands to forested systems. As a product of this initiative, state and transition models such as the one below were produced.
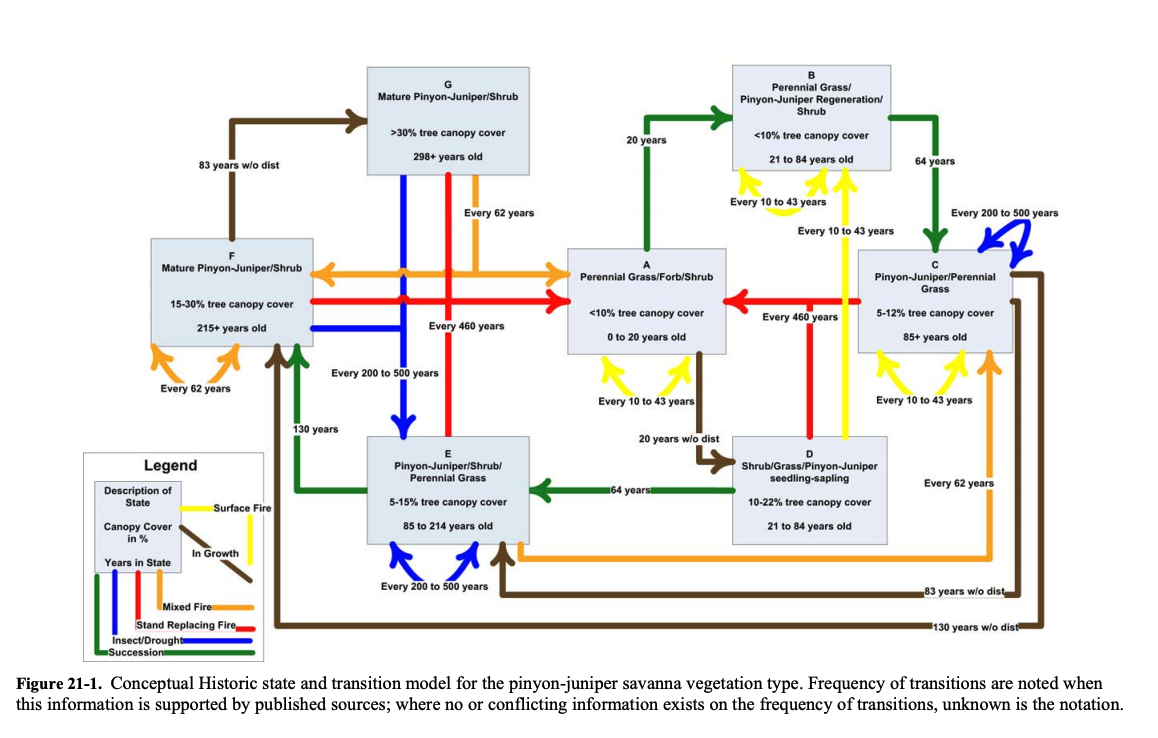
One can read this state-and-transition model (STM) from a number of different perspectives depending on management objectives and the associated desired vegetative community. From an overall rangeland health perspective aimed at an ecosystem that supports biotic integrity, soil stability, and hydrologic function, management objectives may cycle through states A: Perennial Grass/Forb/Shrub, B: Perennial Grass/PJ/Shrub, C: PJ/Perennial Grass, and D: Shrub/Grass/PJ Sapling. If management aims are to maintain habitat for pinyon jays and other species, achieving states with >15 percent tree cover would be the target. A balance among rangeland health attributes and habitat is ideal, however, so the sweet spot of states in the model above may be C: PJ/Perennial Grass (10–12 percent tree cover) and E: PJ/Shrub/Perennial Grass.
Section 5: Current Environmental Anthropogenic Stresses
What is not reflected in the STM above are the current environmental and anthropogenic stresses of livestock grazing, invasive grasses, timber harvest, and mining activities, all of which are covered in the final section of this chapter.
Livestock Grazing
A marked increase in pinyon-juniper tree density and extent over the past century has been attributed by many researchers to a combination of livestock grazing and the disruption of historical wildfire regimes (Gori and Bate 2007; Gottfried et al. 1995). Some have hypothesized that livestock grazing has aided the spread of juniper into grasslands by reducing the density of grasses and palatable shrubs that compete with juniper seedlings for soil moisture, nutrients, and other limiting factors (Johnsen 1962). Livestock overgrazing has modified fire regimes and erosion processes across multiple spatial scales by altering the density and spatial patterning of ground cover (Allen 2007). More trees survive in the absence of fire, further reducing vegetative ground cover by outcompeting grasses and forbs for limited soil moisture. Livestock can also transport nonnative, invasive weed seed from weed-infested rangelands into more pristine areas, where they, too, can outcompete native grasses and forbs.
Erosion
Pinyon-juniper ecosystems in the southwestern United States are particularly susceptible to erosion. When fire, livestock grazing, or drought reduces the amount of vegetative ground cover and organic litter, rates of soil erosion during and after precipitation events can increase dramatically. Where appropriate grazing management systems are not implemented, livestock grazing contributes to soil erosion by compacting the soil. Soil compaction reduces the amount of water that filters down through the soil (infiltration) and increases surface water runoff. As a result, topsoil is lost, and large quantities of sediment are deposited by surface runoff into rivers and streams, negatively impacting water quality and aquatic habitat.
Furthermore, livestock, off-road vehicles, hikers, and other types of mechanical disturbance damage biological soil crusts (communities of bacteria, cyanobacteria, lichens, mosses, algae, and/or fungi on the soil surface). These crusts reduce erosion from wind and water, hold nutrients and moisture, enrich the soil via atmospheric nitrogen fixation, and initiate primary succession by recolonizing disturbed areas (Gottfried et al. 1995). Biological soil crusts play an important role in reducing surface runoff and erosion, but they are fragile and can be slow to recover after disturbance (Belnap and Eldridge 2001).
Invasive Plants
Some invasive plants that cause concern in PJ ecosystems include cheatgrass (Bromus tectorum), knapweeds (Centaurea spp.), thistles (Cirsium spp., Carduus spp., and Onopordum spp.), mustards (Sisymbrium spp. and Brassica spp.), and Dalmatian toadflax (Linaria dalmatica). These species have established and proliferated in PJ systems through vectors such as drought, fire, timber harvest, mining activities, livestock grazing, road construction, and erosion, all of which create conditions that allow nonnative invasive species to become established. Once established, many nonnative invasive plants can outcompete native vegetation for space, moisture, nutrients, and other limiting factors. They spread aggressively and are difficult to control, as they have few natural enemies in their new environment.
Mining Activities
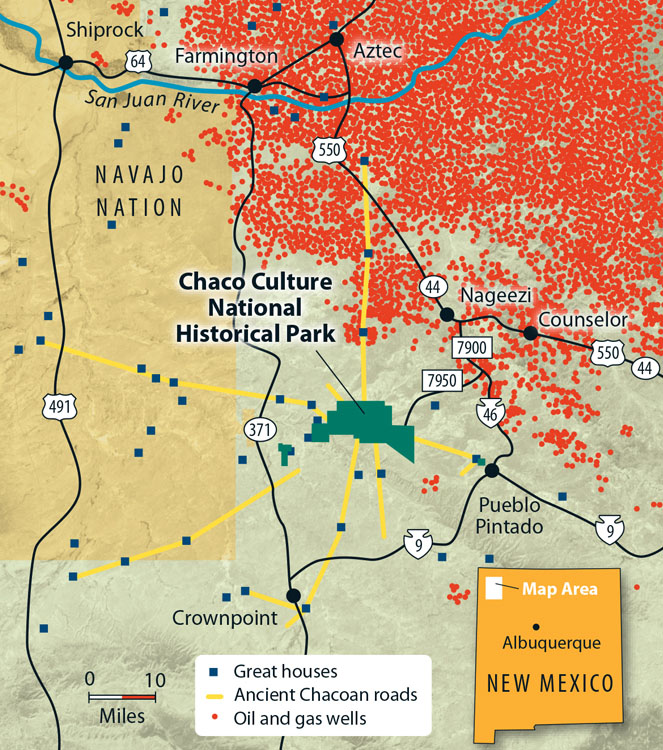
Natural gas extraction is a primary industry in southern Colorado and New Mexico. Maps such as the one shown in figure 9.23 below are typical for large areas throughout the state. This map represents a small portion in the northwest corner of New Mexico, and each red dot is a natural gas well-pad site. There are hundreds of thousands of natural gas well-pad sites throughout PJ ecosystems.
The footprint and ecological impacts of natural gas extraction are severe and far-reaching. Given the number of well-pad sites and associated networks of roads (figure 9.24) simply establishing and maintaining the extraction site is a substantial disturbance. Plant material is removed, and soil is moved to create roads and lay pipeline. This opens niches for invasive species to make their way into the area through natural dispersal and on vehicles coming and going to service equipment. All that traffic compacts and erodes the soil of roads and adjacent areas. During precipitation events, the networks of roads prevent adequate overland water flow.

Further, vegetation clearing fragments and destroys wildlife habitat. This may be through removal of the primary forage or browse plant of a wild herbivore or by altering the plant community structure (height and density) vital for nesting or cover. Some wildlife species require a mosaic of plant types and plant community structures, such as dense shrubs for nesting and open areas for foraging and mating. To some species, such as a small mammal, the width of a road is an uncrossable distance. Disturbed areas become sites for invasive species establishment, and the high degree of vehicle traffic to and from the site serves as a vector of introduction.
